
traduction pour Science & Foi : Benoit Hébert.
Présentation (Benoit Hébert)
En 2009, Alister McGrath était l’orateur des prestigieuses « Gifford Lectures » (voir www.giffordlectures.org/) qui proposent plus de cent ans de réflexion autour de la théologie naturelle.
Alister McGrath a apporté sa contribution sur ce sujet par une série en 6 volets réunis sous le titre « A Fine-Tuned Universe: Science, Theology, and the Quest for Meaning » : « un univers finement réglé : Science, théologie, et la quête de sens ». le programme de chaque conférence est visible sur le site de l’université d’Aberdeen.
Il a alors abordé un thème particulièrement brûlant: la notion de téléologie, c’est à dire de finalité de l’évolution des espèces. L’un des arguments favoris des évolutionnistes athées est de nous faire croire que le mécanisme darwinien serait en lui même la preuve d’une absence de « destin » ou de « but ». Les chrétiens qui pensent que l’évolution est la méthode choisie par Dieu pour créer les espèces vivantes ont différentes façons de répondre à cette question. La plupart du temps, ils se contentent d’affirmer que la notion de « téléologie » est une notion métaphysique que l’on ne peut pas tester scientifiquement et qui échappe donc au cadre de la science. Ainsi, celui qui dans l’histoire de la vie (comme dans l’histoire tout court d’ailleurs, où la notion de contingence est évidente) voit la main de Dieu met sa foi en action. Celui qui refuse d’y voir l’action divine met lui aussi sa conviction athée en jeu, ainsi chacun « vit par la foi ».
Alister McGrath se place aussi sur le terrain de la science:
« Ainsi, existe-t-il une direction de l’évolution, que l’on choisisse de l’interpréter de manière téléologique ou pas? La manière de poser la question montre bien qu’il s’agit là d’une question légitime scientifiquement, et pas d’une question théologique spéculative. La vision d’une évolution indéterminée en terme d’aboutissement, sans prédiction possible a fini de dominer la biologie évolutive. »

Les énigmes de la biologie de l’évolution
Dans ma conférence précédente, j’ai abordé la notion passionnante et controversée du « réglage minutieux » de l’univers, en particulier la valeur de certaines constantes fondamentales de la physique. Pourtant, cette notion de « réglage minutieux » n’est pas cantonnée au domaine de la cosmologie, on en parle aussi en biologie. Son interprétation soulève des difficultés importantes qu’il nous faut confronter. L’un des axiomes de base du darwinisme est que la nature est capable de se régler elle-même au travers de mécanismes évolutifs, même si ce réglage ne produira pas la meilleure solution, mais plutôt une solution fonctionnelle et viable. Même si on arrivait à montrer qu’il existe un « réglage minutieux » dans les systèmes biologiques évolués, beaucoup argumenteraient que de telles observations s’expliquent sans problème dans le cadre de l’orthodoxie darwinienne. L’exemple de l’œil humain est un exemple excellent, c’est un système complexe pour lequel un mécanisme évolutif entièrement plausible peut être proposé, contrairement à la vision statique de William Paley.
Pourtant, le concept de « réglage minutieux » ne perd rien de son pouvoir à stimuler notre réflexion. Comme le théologien Charles Kingsley (1819-75) l’a fait remarquer en 1871 dans sa conférence « La théologie naturelle du futur », les théologies naturelles anciennes, y compris celle de Paley, était basées sur la croyance que Dieu fabriquait toute chose- alors qu’une théologie naturelle moderne nous décrit un Dieu « bien plus sage que cela », en ce que Dieu a choisi « de faire toutes choses se faire elles-mêmes. » La vision statique de Paley a été remplacée par une conception de la providence et de la causalité dans laquelle l’action et la présence permanente de Dieu sont affirmées au sein de l’ordre naturel évoluant lentement. Ainsi, l’ordre créé est tel qu’il possède une capacité « interne » à évoluer, grâce à laquelle des structures nouvelles « émergent » véritablement.
Cette nouvelle approche de la théologie naturelle n’est qu’un épisode dans la longue liste des reformulations du contenu conceptuel fondamental permettant de répondre au besoin religieux des différentes périodes historiques et culturelles- dans ce cas, à l’occasion de la publication de l’Origine des Espèces de Darwin en 1859 et alors que la vision statique et rigide de Paley de la création engendrait une frustration grandissante. Cette nouvelle approche due à Kingsley entrait en résonance avec la culture émergente… Même le « bulldog de Darwin », Thomas Henry Huxley, concéda qu’il n’y avait aucune raison de principe qui empêcherait le mécanisme évolutif d’être inclus dans une conception initiale de l’univers.
À un certain moment, les mécanismes nécessaires à l’évolution ont émergé, nous ne comprenons pas encore clairement comment. Néanmoins, il est clair que la capacité à coder l’information est vitale pour l’évolution en général, et la capacité à évoluer en particulier. Et ceci est directement lié à la chimie du carbone, qui permet la formation de longues chaînes stables. Aucun autre élément ne le permet ; sans cela, pas d’ARN, ni d’ADN, ni de mécanismes de duplication associés. La capacité de l’évolution à s’autoréguler est donc dépendante ultimement des propriétés chimiques du carbone, qui sont elles mêmes un exemple solide de réglage minutieux.
On néglige bien souvent cet aspect des choses dans la description de l’évolution, en considérant que la chimie et la physique ne sont qu’un support nécessaire mais sans grand intérêt dans une discussion à propos de
l’évolution. Pourtant, ce processus biologique requiert une planète stable, irradiée par une source d’énergie capable de conversion chimique et de stockage, et l’existence de divers éléments chimiques, avec certaines propriétés fondamentales, avant même que la vie puisse éclore, sans même parler d’évoluer. On a tellement été familier en biologie à l’existence d’agrégats très organisés qu’on ne porte plus aucune attention aux conditions qui ont rendu possible leur existence. On suppose implicitement que la vie ce serait adaptée à tous les milieux, quelles qu’aient été leurs caractéristiques physiques ou chimiques. Rien pourtant ne le prouve et c’est très discutable. L’un des arguments centraux de notre propos est que l’émergence de la vie ne peut pas être étudiée isolément de l’environnement qui a créé les conditions et a fourni les ressources qui l’ont rendue possible.
On reconnaît généralement que les systèmes vivants présentent deux caractéristiques essentielles : un système métabolique autonome, un système génétique capable de transmettre l’information biologique. Pourtant, il est surprenant qu’on accorde si peu d’attention aux propriétés chimiques nécessaires à ces deux processus. Et si la chimie terrestre avait été différente ? Ces dernières années, on a pris conscience des contraintes imposées par les contraintes chimiques à l’évolution. Tout en reconnaissant pleinement le rôle de la contingence dans les processus évolutifs, certains chercheurs ont insisté sur le fait que l’évolution était nourrie, guidée et contrainte par les changements chimiques de l’environnement, avec beaucoup de conséquences inévitables. De tels argument doivent être replacés dans le contexte plus large d’un changement conceptuel en biologie évolutive, dans lequel le caractère aléatoire traditionnel du néodarwinisme est entrain d’être supplanté par une émergence de la vie davantage régulée par des lois scientifiques. Les principes fondamentaux de la thermodynamique et de la chimie confèrent à l’évolution une direction forte. L’évolution n’est plus vue comme un processus purement aléatoire, mais elle apparaît avoir été guidée dans une progression prévisible à partir d’organismes unicellulaires vers des plantes et des animaux sous l’effet de contraintes chimiques.
L’évolution a-t-elle un « but »?
L’un des débats actuels les plus important en philosophie de la biologie est de savoir si on peut considérer que l’évolution est téléologique (c’est-à-dire qu’elle a un but). Le rejet de toute forme de téléologie a acquis le statut de vérité axiomatique dans le néodarwinisme depuis les années 1970. On considérait alors que l’évolution était un processus ouvert et indéterminé, sans aucun but prédéfini. Cette position a émergé très tôt dans la réception populaire de la théorie de la sélection naturelle de Darwin. Comme on le souligne souvent, ce qui a frappé Thomas H. Huxley dans sa première lecture de L’Origine des Espèces était « la conviction que la téléologie, telle qu’on la concevait habituellement, avait reçu le coup de grâce des mains de Mr Darwin. » Cette conception de la téléologie était présente dans la Théologie Naturelle (1802) de Paley, qui soulignait que la nature était conçue et construite avec une intention et un but spécifique à l’esprit.
Pourtant, nous devons nous demander si certains darwinistes ne font pas exactement le même genre de spéculation métaphysique, ou s’ils ne sont pas piégés malgré eux par le même type de préjugés métaphysiques, que ceux qu’ils identifient chez ceux qui ont une approche téléologique de la biologie. L’appel grandissant au darwinisme chez ceux qui souhaitent renverser l’approche de Paley en développent une athéologie naturelle est très important en ce que cela nous montre qu’une supposition de travail en biologie a été transposé en un dogme de l’athéisme fondamentaliste. Le biologiste de Princeton C. S. Pittendrigh (1918-96) a introduit le terme « téléonomie » en 1958 en biologie … Ce concept a été développé plus tard par Jacques Monod ( 1910-1976), qui a affirmé que la téléonomie avait remplacé la téléologie en biologie évolutive. Ainsi, Monod souhaitait montrer que la biologie évolutive avait comme objectif d’identifier et de clarifier les mécanismes impliqués dans le processus évolutif….Ces processus n’avaient pas d’objectif. On ne pouvait donc pas parler de « but » de l’évolution.
Cette explication de Monod et son emphase sur les évènements « aléatoires » nous donne l’exemple d’une tendance générale en biologie de l’évolution dans l’importance qu’elle donne aux approches statistiques. On peut faire remonter cette tendance à R. A. Fisher, Theodosius Dobzhansky et J. B. S. Haldane. Pourtant, comme Phillip Sloan l’a fait remarquer, les modèles mathématiques idéalisés de ces réinterprétations de la théorie de la sélection naturelle impliquaient l’incorporation de processus stochastiques et aléatoires à un niveau théorique. L’emphase sur le caractère « sans but » de la sélection naturel que l’on retrouve chez beaucoup d’auteurs néo darwiniens, n’est rien de plus qu’un « ornement rhétorique injustifié », qui s’appuie sur la « réification de ces modèles idéalisés de dynamique des populations en affirmations réalistes et métaphysiques sur le monde. » Dans un cadre théologique réformé, « aléatoire » peut être traduit par « non prédictible », et peut ainsi être placé dans la doctrine générale de la providence divine. De plus, que l’évolution nous démontre ou pas l’existence d’une conception, une intention ou un but, elle démontre sans équivoque une direction. Les organismes sont généralement devenus plus gros, plus complexes, plus diversifiés d’un point de vue taxonomique…Cela implique-t-il une téléologie ? Pour répondre à cette question, il nous faut préciser ce que le terme « téléologie » signifie. Il y a de bonnes raisons de penser que l’utilisation de ce mot est légitime en biologie, au moins dans une certaine mesure. La notion de « programme génétique » développée par François Jacob et d’autres dans les années 1970 peut être vue comme une validation partielle de cette notion. Comme Jacob l’a dit, la téléologie est comme une « maîtresse »- quelqu’un dont les biologistes ne peuvent pas se passer, mais avec laquelle ils ne veulent pas être vus en public. » Francisco Ayala affirme que la notion d’explication téléologique est fondamentale en biologie moderne. Elle est nécessaire pour rendre compte des rôles fonctionnels habituels des différentes parties des organismes vivants, ainsi que pour décrire le but de l’aptitude reproductive qui joue un rôle essentiel dans la sélection naturelle.
Une explication téléologique sous entend que le système considéré est organisé dans une direction. Pour cette raison, les explications téléologiques sont appropriées en biologie et en cybernétique, mais n’ont aucun sens dans la description des phénomènes tels que la chute d’une pierre. Plus essentiel encore, les explications téléologiques impliquent que le résultat final est la raison explicative de l’existence de l’objet ou du processus qui y conduit. Une explication téléologique des branchies des poissons implique que ces branchies sont apparues précisément parce qu’elles permettent la respiration. Si le raisonnement ci-dessus est correct, l’utilisation d’explications téléologiques en biologie est non seulement acceptable mais aussi indispensable. Pour Ayala, la sélection naturelle elle-même, source ultime d’explication en biologie évolutive, est ainsi un processus téléologique à la fois parce qu’elle dirigée vers une augmentation de l’efficacité de la reproduction et parce qu’elle produit les organes et les processus dirigés pour ce but. Les mécanismes téléologiques dans les organismes vivants sont donc des adaptations biologiques, qui ont surgi comme résultats du processus de la sélection naturelle.
Peut-on parler de « direction » de l’évolution?

Ernst Mayr (1904-2005), que l’ont crédite largement de l’invention de la philosophie moderne de la biologie, tout spécialement de la biologie évolutive, présente quatre objections traditionnelles à l’utilisation du langage téléologique en biologie.
1Les affirmations ou les explications téléologiques impliquent l’adhésion à des suppositions d’ordre métaphysique ou théologique invérifiables dans le cadre de la science. Mayr a par exemple l’exemple de l’élan vital de Bergson ou les écrits de Hans Driesch (1867-1941).
2La croyance que l’acceptation d’explications de phénomènes biologiques qui ne seraient pas applicables à la nature inanimée constitue un rejet d’explication physico-chimique.
3La supposition que des objectifs futurs sont la cause d’évènement actuels parait incompatible avec la notion courante de causalité.
4Le langage téléologique semble être un anthropomorphisme très discutable. L’utilisation de mots tels que « dirigé pour un but » parait être un transfert de qualités humaines- tels que la planification et la recherche d’objectifs- à des structures organiques.
Comme Mayr le souligne, le résultat de ces objections et d’autres a conduit au fait que les explications téléologiques en biologie ont été couramment considérées comme une « forme d’obscurantisme ». Pourtant, les biologistes continuent d’utiliser un langage téléologique, en insistant sur son utilité et sa pertinence méthodologique et heuristique. De sérieuses objections ont été et sont encore élevées à propos d’une conception de l’évolution considérée comme un agent conscient, planifiant activement ses objectifs et ses résultats, ou bien conduite vers un objectif prédéfini par une force mystérieuse.
Pourtant, il faut noter que de telles façons anthropomorphiques de parler (et de penser) sont évidentes dans certaines sections de la biologie contemporaine. Un parfait exemple nous est fourni par la description évolutive du « gène de l’œil », popularisée par Richard Dawkins, qui décrit le gène comme un agent actif. Tout en avertissant soigneusement que « nous ne devons pas concevoir le gène comme un agent actif et intentionné. », Dawkins souligne que le processus de la sélection naturelle « les fait plutôt agir comme s’ils avaient un objectif ». Cette façon anthropomorphique de parler sous entend l’attribution d’une intentionnalité et d’une activité à une entité qui n’est en définitive qu’un participant passif dans le processus de réplication, plutôt que son agent actif. Plus sérieusement, en affirmant que le gène est un répliquant, Dawkins paraît lui conférer une certaine forme d’autonomie biologique, ce qui met sous silence sa position au sein d’un système.
Bien entendu, l’émergence de cette approche du « gène de l’œil » doit être contextualisée et envisagée à la lumière du rejet de formes plus élevées de sélection naturelle décrite dans le livre influent de George C. Williams Adaptation and Natural Selection (1966). La critique de Williams à propos de la théorie de la sélection de groupe a été le début d’un paradigme nouveau en matière de sélection des gènes, en affirmant que la sélection naturelle est principalement, sinon exclusivement, une sélection en faveur ou en défaveur de gènes uniques. Dans Le gène égoïste (The Selfish Gene (1976)), Dawkins a largement popularisé cette approche et a contribué à son acceptation. Pourtant, aujourd’hui le vent a tourné, et les approches hiérarchiques ou à plusieurs niveaux de la sélection naturelle ont regagné du terrain chez les biologistes.
Comme Mayr le souligne à juste titre, la nature abonde en processus et en activités qui conduisent vers un objectif. Cependant, nous choisissons de les interpréter. Les exemples de comportements dirigés vers un but sont très répandus dans le monde naturel ; en effet, « l’existence de processus dirigés vers un but est peut-être l’un des traits les plus caractéristiques des systèmes vivants. » La reformulation d’affirmations téléologiques sous une forme non téléologique conduit invariablement à des « platitudes sans signification ». Bien qu’il entoure sa conclusion de bien des précautions, Mayr insiste sur le fait qu’il est approprié de dire que : « l’usage de ce que l’on appelle le langage « téléologique » est légitime en biologie ; cela n’implique pas le rejet d’explications physico chimiques pas plus que d’explications non causales. »
Un autre problème surgit de la notion de “capacité à évoluer”. Certains affirment que la nature semble sélectionner préférentiellement les formes capables de développements évolutifs futurs. Marc Kirschner et John Gerhart donne une définition utile de cette notion dans une étude remarquable de 1998 : « La capacité d’une lignée à évoluer est aussi qualifiée de capacité d’adaptation évolutive. Il s’agit de sa capacité à générer des variations héritables et sélectionnables de son phénotype. » Pourtant, ceci semble bien lier la sélection avec les caractéristiques d’états futurs- ce qui contredit clairement la troisième objection de Mayr, qui rejette toute possibilité que « des objectifs futurs soient la cause d’évènement actuels », ce qui contredirait les notions courantes de causalité.
Ainsi, existe-t-il une direction de l’évolution, que l’on choisisse de l’interpréter de manière téléologique ou pas? La manière de poser la question montre bien qu’il s’agit là d’une question légitime scientifiquement, et pas d’une question théologique spéculative. La vision d’une évolution indéterminée en terme d’aboutissement, sans prédiction possible a fini de dominer la biologie évolutive. Beaucoup d’auteurs ont adopté le paradigme darwinien standard affirmant le caractère essentiellement aléatoire et contingent de la nature dans les processus évolutifs.
Ainsi, existe-t-il une direction de l’évolution, que l’on choisisse de l’interpréter de manière téléologique ou pas? La manière de poser la question montre bien qu’il s’agit là d’une question légitime scientifiquement, et pas d’une question théologique spéculative. La vision d’une évolution indéterminée en terme d’aboutissement, sans prédiction possible a fini de dominer la biologie évolutive. Beaucoup d’auteurs ont adopter le paradigme darwinien standard affirmant le caractère essentiellement aléatoire et contingent de la nature dans les processus évolutifs. Par exemple, Stephen Jay Gould (1941-2002) insistait sur le fait que “pratiquement tous les évènements intéressants de l’histoire de la vie peuvent s’interpréter en terme de contingence ». Il est inutile de parler de but, d’inévitabilité historique ou de direction. Du début jusqu’à la fin, le processus évolutif est dirigé par les contingences. »Nous sommes le résultat accidentel d’un processus non planifié… le résultat fragile d’une accumulation énorme d’évènements improbables ; et non le résultat d’un processus défini. » Comme le Gould le raconte en utilisant cette analogie célèbre des années 1990, si nous devions repasser la K7 vidéo du film de la vie, nous verrions quelque chose de différent à chaque fois. « Repasser le film une autre fois et la première étape des cellules procaryotes vers les cellules eucaryotes pourrait bien durer 12 milliards d’années au lieu de deux. » L’influence de la contingence est telle que tout ce qui est arrive est le fruit des coïncidences. « Modifier tout événement du passé, même de manière insignifiante, et l’évolution se déroulera de façon radicalement différente. » Gould affirme que la contingence est si essentielle que la K7 montrera un film différent à chaque fois qu’elle sera visionnée. Ainsi, le processus du développement évolutif est-il réellement aussi dépendant des coïncidences de l’histoire ?

Contingence ou convergence ?
L’emphase de Gould à propos de la contingence historique est envisagée avec beaucoup de suspicion au sein même de la communauté des biologistes de l’évolution. Par exemple, Leigh van Valen critique l’utilisation de la métaphore de la « K7 vidéo de la vie » faite par Gould. Qu’arriverait-il vraiment, demande van Valen, si comme le suggère Gould nous repassions le film de la vie une nouvelle fois ? Van Valen concède immédiatement qu’il y aurait à coup sûr des différences entre ces deux versions de la K7. Toutefois, par un examen plus attentif, on verrait que la situation serait bien plus complexe que ce que Gould affirme. Malgré les différences, il y aurait aussi des ressemblances. Repasser le film de la vie encore quelques autres fois. Nous verrons des motifs mélodiques similaires apparaître dans chaque, et la structure d’ensemble pourrait être tout à fait similaire…Si nous observons la situation d’un point de vue plus large, le rôle de la contingence diminue. Regardez la K7 comme un tout. Elle ressemble par certains aspects à une symphonie, bien que son orchestration soit interne et largement provoquée par les interactions entre beaucoup de lignes mélodiques. Bien que les détails puissent être différents, van Valen soutient que l’on doit s’attendre à des similarités et des convergences.
Une approche similaire est adoptée par le paléo biologiste Simon Conway Morris, dont le travail pionnier sur le site schisteux de Burgess a été utilisé par Gould, dans des façons que Conway Morris désapprouvait clairement. Bien que Gould et Conway Morris reconnaissent tous deux le rôle de la contingence dans les processus évolutifs, ils en évaluent très différemment l’importance. Pour Gould, l’homme est le résultat de la contingence de l’évolution adaptative : « la formidable improbabilité de l’évolution humaine ». Conway Morris affirme que si notre planète était un tant soit peu différente de ce qu’elle est effectivement, alors la vie n’aurait jamais émergée. Bien que ceci puisse sembler comparable à l’emphase de Gould à propos de la contingence, il est important de noter que Conway Morris souligne la façon dont les événements physiques créent des occasions propices à l’émergence de la vie et à son adaptation. Conway Morris caractérise son travail comme la réfutation de la « notion de domination de la contingence ». Dans Life’s Solution, Conway Morris affirme qu’il n’existe qu’un nombre limité de points d’arrivée au processus évolutif.
« Repasser le film de la vie aussi souvent que vous voudrez, et la fin du film sera presque la même. »
Ce livre souligne le caractère prédictible des aboutissements du phénomène évolutif, pas en terme de détails génétiques mais plutôt dans leurs manifestations phénotypiques plus large. L’évolution convergente doit être envisagée comme :
« la tendance récurrente de l’organisation biologique d’arriver aux mêmes solutions répondant à un même besoin particulier. »
La démonstration de Conway Morris est basée sur une accumulation remarquable d’exemples d’évolution convergente, dans lesquels deux ou plus de lignées ont évolué indépendamment vers des structures et des fonctions similaires. Les exemples cités par Conway Morris vont de l’aérodynamisme des papillons de nuits et des oiseaux mouches, à l’utilisation de toiles par les araignées et d’insectes pour capturer leurs proies. « Les détails de la convergence nous montrent effectivement bien des tours et détours lors des changements évolutifs qui fournissent autant de points de départs et sont transformés en solutions communes via une variété de sentiers tous tracés.” Et quelle est la signification de l’évolution convergente ? Conway Morris est clair : cela révèle l’existence de région stable dans l’espace biologique. « La convergence se produit parce qu’il existe des « îles de stabilité », analogues aux attracteurs de la théorie du chaos. »
On ne peut sous estimer la critique que Morris fait de Gould. Alors que la contingence est un facteur des mécanismes évolutifs, son rôle est bien moins décisif que Gould ne l’affirme. L‘évolution paraît converger régulièrement vers un nombre relativement petit d’aboutissements possibles. La convergence est partout, en dépit de l’infinité des possibilités génétiques parce que :
Les routes de l’évolution sont nombreuses, mais les destinations sont limitées.
Certaines destinations évolutives sont vouées à l’extinction, car certains des phénotypes mal adaptés ne sont pas viables, supprimant ainsi toute exploration future par la sélection naturelle. L’histoire de la vie nous montre une tendance à la répétition, la vie nous démontre son inlassable habileté à se frayer un chemin jusqu’à la bonne solution.
La vie a une tendance particulière à naviguer vers des solutions précises en réponse à des défis adaptatifs.
L’importance de la convergence évolutive
Les exemples d’évolution convergente sont légion.
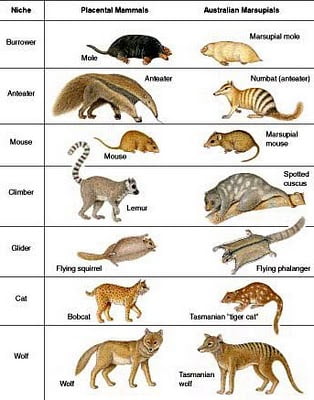
Les mammifères placentaires et les marsupiaux (Australie) ont évolué de manière indépendante. Ils se ressemblent pourtant énormément car ils remplissent les mêmes niches écologiques.
Pour ceux qui ne seraient pas familiers avec cette idée, on peut donner brièvement deux exemples.
1La photosynthèse. On connaît trois mécanismes différents, que l’on appelle couramment « métabolisme de l’acide Crassulacéen » (CAM), C-3 et C-4. La photosynthèse de type C-4 a évolué au moins 31 fois dans 18 familles différentes de plantes à fleurs durant les 8 millions d’années précédents, donnant naissance à un total de près de 10 000 espèces de plantes. Le mécanisme CAM a lui aussi évolué à plusieurs occasions.
2L’œil. L’évolution de l’œil a connu un développement spectaculaire lors de l’explosion cambrienne. Ce processus a combiné une variabilité morphologique remarquable avec une stase génétique et du développement pendant des millions d’années d’évolution. Les yeux ont évolué à de multiples occasions indépendantes, en prenant au moins neuf formes différentes : des yeux « trous d’épingles », deux sortes d’ »yeux caméra », des yeux « lentilles » (chez les vertébrés et les pieuvres), des yeux avec des « réflecteurs incurvés », ainsi qu’avec différents composants, des yeux aux « lentilles multiples ». Les yeux composés ont évolué de manière indépendante chez les crustacés, Les vers annélides, et les mollusques bivalves. Les « yeux caméra » n’ont pas seulement évolué chez les vertébrés et les pieuvres, mais indépendamment chez les araignées sauteuses, chez certains escargots, chez certains vers (alciopes polychaete), la méduse cubozoénne.
Voici ce que Morris espère montrer en rassemblant ces exemples de convergence: le nombre d’aboutissements possibles est limité. Encore et encore, l’évolution « converge » vers un nombre relativement petit de solutions possibles aux problèmes et aux opportunités qu’offre l’environnement à la vie. Ainsi, même si le mécanisme de recherche est essentiellement aléatoire, celui-ci aboutira en identifiant des solutions stables dans l’espace biologique. Alors que la façon de trouver ces « îlots de stabilité » peut sembler erratique, son aboutissement est complètement intelligible.
Gould suggère que la direction de l’évolution pourrait être comparée à la « démarche d’un homme ivre », dans laquelle les organismes errent vers une plus grande complexité. Conway Morris propose une alternative à la dichotomie rigide proposée si souvent entre le caractère purement aléatoire (la marche au hasard de Gould) et une progression étroitement dirigée vers un but final pré-établi. En faisant et en défendant ce point de vue important, Conway Morris propose une analogie non biologique très parlante. Il raconte la découverte de l’île de Pâques par les Polynésiens, il y a peut-être 1200 ans. L’île de Pâques est l’un des endroits les plus reculés de la terre, à au moins 3000 km des centres de population humaine les plus proches : Tahiti et le Chili. Bien qu’entouré par les vastes étendues de l’océan Pacifique, elle a toutefois été découverte par les Polynésiens. Peut-on attribuer ceci au hasard ou à une coïncidence ? demande Morris. C’est possible, mais probablement pas. Conway Morris relate alors la « stratégie de recherche sophistiquée des Polynésiens » qui a rendu cette découverte inévitable. Il affirme que la même chose se produit dans les processus évolutifs : « les îles isolées fournissent des havres de possibiltés biologiques dans un océan de ‘mauvaise adaptation’ » Ce sont ces îles de stabilité qui donnent naissance au phénomène d’évolution convergente. Peut-on prévoir ces « îles de stabilité » ? Peut-on identifier à l’avance, pour ainsi dire les points cers lesquels le processus évolutif va converger ? Conway Morris est très prudent sur cette question. Après tout, la méthode scientifique concerne des analyses a posteriori, pas des prédictions a priori. “La sagesse prédictive est strictement interdite. . . nous ne pouvons pas prévoir mais seulement constatera a posteriori.” La théorie de l’évolution fournit sûrement un compte rendu de ce que l’on a observe – mais elle ne peut pas prévoir les spécificités du future. La notion d’îles de stabilité est toutefois parfaitement valide, et peut être appliquée rétroactivement sur la base de ce que nous savons déjà des paramètres probablement impliqués dans le processus évolutif. Il est possible que les îles de stabilité ne puissent être prédites qu’avec difficulté, il est pourtant possible de prédire le phénomène global, et l’identité de ces « îles » rétroactivement, sur la base de cette compréhension des forces contingentes.
Ainsi, parce que les organismes parviennent de façon répétée aux mêmes solutions biologiques-l’œil caméra des vertébrés et celui des céphalopodes en sont un parfait exemple- cela suggère non seulement qu’il y a un degré de prédictibilité au processus évolutif, mais ceci pointe potentiellement vers une structure plus profonde de la vie, un « paysage métaphorique à travers lequel l’évolution doit obligatoirement naviguer. »
Où ces quelques lignes nous conduisent-elles donc ? Il est clair que l’analyse de Conway Morris pointe vers une forme de téléologie- pourtant, ce concept peut facilement être incorporé dans le spectre des possibilités utilisées habituellement dans ce genre de discussion.
Il nous faut maintenant faire deux remarques fondamentales. Premièrement, la plupart des objections faites au recours à la notion de téléologie en biologie et relevées par Mayr reflètent la croyance qu’un système métaphysique (souvent théiste) est imposé a priori sur le processus d’observation scientifique et à la réflexion, étant ainsi préjudiciable à la démarche scientifique. Une lecture attentive de Mayr suggère qu’il pense –non sans quelques bonnes raisons- que la notion spécifique de téléologie de Kant a exercé un effet généralement négatif sur le développement de la philosophie de la biologie. L’origine et l’influence de la notion kantienne de téléologie a été l’objet d’intenses investigations dans les dernières années, validant (mais pas totalement) les préoccupations de Mayr à propos de l’intrusion de notions métaphysiques a priori dans la démarche scientifique.
Du point de vue de la méthode scientifique, on pourrait en effet protester contre le fait d’imposer a priori des notions de buts ou de causes, telles que celles associées aux nombreuses approches traditionnelles de la téléologie. La même suspicion intense de notions métaphysiques est associées à la montée du « positivisme logique » au cours du vingtième siècle. Toutefois, l’échec ultime d’une attitude empirique aussi radicale était fondée sur son rejet a priori d’une possibilité a posteriori de principes ou d’entités métaphysiques. Le même raisonnement s’applique en matière de téléologie. Les sciences naturelles ont bien raison de protester contre l’intrusion de schémas téléologiques préconçus dans l’analyse scientifique. Mais qu’en est-il si de tels schémas émergent des processus de réflexion à propos des observations ? Qu’en est-il s’il s’agit de conclusions a posteriori plutôt que des suppositions a priori ? Les indices et l’analyse de Conway Morris suggèrent qu’une forme de téléologie peut en effet être déduite a posteriori, en tant que « meilleure explication » de ce que l’on observe. Ceci ne pointera pas peut-être pas directement avec la doctrine traditionnelle de la providence ; néanmoins, il existe un degré significatif de résonance avec cette dernière notion qui mérite une attention plus particulière.
Evolution et théologie naturelle
 Il faut noter qu’il ne s’agit pas nécessairement de discerner un « but » de l’évolution- une notion lourdement connotée d’un point de vue métaphysique- et de prouver ainsi que Dieu a ordonné le processus. Notre approche ressemble davantage à celle du théologien John Henry Newman faisant la remarque étonnement peu étudiée :
Il faut noter qu’il ne s’agit pas nécessairement de discerner un « but » de l’évolution- une notion lourdement connotée d’un point de vue métaphysique- et de prouver ainsi que Dieu a ordonné le processus. Notre approche ressemble davantage à celle du théologien John Henry Newman faisant la remarque étonnement peu étudiée :
Je crois dans une conception (design) de l’univers parce que je crois en Dieu ; pas en Dieu parce que je vois une conception (design).
En accord avec l’approche générale de ces conférences, nous nous posons une question plus « oblique » : le processus évolutif, en dépit de son caractère contingent, ne pourrait-il cependant pas être consonant avec l’accomplissement du but fixé par un Dieu créateur ?
La théorie de Darwin indique certainement qu’il n’est plus nécessaire de faire appel à un Dieu créateur pour rendre compte de la conception apparente des êtres vivants, dans la mesure ou l’on peut argumenter que cette conception est le fruit d’une interaction complexe entre le hasard et la nécessité, entre son caractère aléatoire et des processus déterministes, dans le processus de sélection naturelle. Pourtant, alors que ceci démontre qu’une justification théiste de la conception biologique n’est pas nécessaire, cela ne démontre pas l’affirmation bien plus forte et lourde de signification que la foi en un Dieu personnel (le théisme) et créateur est fausse. Ainsi, les théistes sont libres de penser que les processus naturels sont adéquats pour expliquer la conception des êtres vivants, mais ils sont aussi libres pour insister sur le fait que le théisme leur fournit une explication plausible et rationnelle qui pourrait ultimement être vue comme la meilleure explication.
Encore une fois, le problème concerne la consonance ou résonance entre une vision chrétienne du monde et ce que nous observons effectivement. Les découvertes téléonomiques de la biologie de l’évolution, toutefois limitées, peuvent être facilement réconciliées avec une vision chrétienne de la réalité. Et, comme nous l’avons déjà souligné, la notion de “création” n’exige pas d’être interprétée comme un unique événement, ayant eu lieu une fois pour toute, mais peut tout aussi bien – et beaucoup diraient justement- être comprise comme un processus dirigé. Il nous faut répéter ici la citation de Charles Kingsley : « Nous savions que Dieu était si sage qu’il pouvait faire toute chose : mais bien plus, il est tellement plus sage que cela qu’il peut faire que toutes choses se fassent d’elles mêmes. »
L’approche que nous avons soulignée dans cette conférence étend le concept de théologie naturelle des aboutissements des processus évolutifs à une appréciation des processus par eux-mêmes. La vision de la nature essentiellement statique de Paley focalisait inévitablement sont attention sur l’état présent des choses. Notre approche, sans pour autant diminuer en quoi que ce soit la beauté et l’émerveillement du monde naturel autour de nous, étend ce sens d’émerveillement aux processus qui y ont abouti. Les processus et leur aboutissement sont tous deux objets d’une théologie naturelle.
Ernst Mayr et d’autres philosophes ont sûrement raisons de protester contre toute tentatives d’imposer une téléologie prédéterminée sur le compte rendu scientifique du processus évolutif. Pourtant, les arguments de Mayr n’ont de force réelle que contre les concepts téléologiques a priori, qui sont importés de systèmes métaphysiques non empiriques, que ce soient des systèmes théistes ou athées. J’ai argumenté qu’une certaine notion de téléologie émerge de l’étude des processus évolutif eux-mêmes. Une telle téléologie est empirique, fondée sur un discernement a posteriori, pas une supposition a priori. Elle est déduite d’une observation des processus évolutifs, non déduite d’un système métaphysique non empirique. Le terme « téléologie » est plus élastique que ses critiques ne semblent le réaliser. Il faut modifier sa signification à la lumière des données empiriques, mais pas l’abandonner en réponse aux demandes dogmatiques de ceux qui maintiennent son inconcevabilité.
Tous ces points vont en faveur du caractère non éradiquable du langage et de la pensée téléologique en biologie. Il est facile d’en faire le portrait d’une manière obsolète de s’exprimer qui disparaîtra avec le temps et l’éducation rigoureuse de la méthode scientifique. Pourtant, ce jugement est superficiel et inutile. La pensée téléologique persiste en biologie précisément parce qu’elle apporte du sens aux observations, ce qui résonne avec une façon “naturelle” et humaine de penser. De la même façon que l’on parle d’un gène « égoïste », on peut parler de l’évolution comme allant vers « un but ». Les deux expressions sont des manières anthropomorphiques de parler ; pourtant, toutes deux peuvent apporter un éclairage valide.
Conclusion
La biologie de l’évolution doit à présent expliquer la navigation apparente du processus de recherche évolutif vers des régions stables de l’espace biologique. Il est difficile de voir comment un langage téléologique, même minimaliste, pourrait être évité. Comme Conway Morris le suggère, en utilisant l’image du « compas de Darwin » : La vision d’une évolution avec une fin indéterminée, sans prédiction possible et indéterminée en terme d’aboutissement est contredite par l’omniprésence de la convergence de l’évolution. L’argument central est que, puisque les organismes parviennent de façon répétée aux même solutions biologiques, l’œil « caméra » des vertébrés et des céphalopodes en est peut-être l’exemple le plus connu, cela confère non seulement une possibilité de prédiction mais cela indique de manière intrigante qu’il existe une structure plus profonde à la vie, un paysage au travers duquel l’évolution doit obligatoirement naviguer.
Peut-être que certains craignent que nous introduisions arbitrairement des notions métaphysiques controversées comme celles de but ou de téléologie dans une discussion scientifique neutre. C’est une crainte justifiée. Même une lecture rapide des travaux contemporains en biologie de l’évolution nous montre comment des agendas théologiques ou anti- théologiques s’introduisent fréquemment dans ce qui est supposé être une discussion scientifique neutre et objective. Ce qui est présenté comme une analyse empirique de la réalité se révèle souvent être infesté de suppositions non empiriques, couvrant souvent des dogmes métaphysiques.
Pour illustrer ce dernier point, nous pourrions considérer une affirmation faite par Richard Dawkins, zoologiste d’Oxford dans le Gène Egoïste, publié en 1976. Dawkins illustre ici un point de vue fréquent concernant la vision du « gène de l’œil » de l’évolution, qui était le point de vue dominant dans les cercles évolutionnistes à cette époque, dans lequel le gène est décrit comme un agent actif :
Les gènes grouillent dans de grandes colonies, en sécurité à l’intérieur de robots massifs, sans possibilité d’aller dans le monde extérieur, communicant avec lui par des moyens indirects tortueux, le manipulant par un contrôle à distance. Ils sont en vous en moi ; ils nous ont créé, corps et esprit ; et leur conservation est la raison ultime de notre existence.
Dawkins fait ici une affirmation empirique- les gènes « sont en vous en moi »- et l’entoure d’un ensemble d’interprétations, que le lecteur non averti pourrait interpréter comme des observations directes, sans se rendre compte à quel point elles sont connotées d’un point de vue métaphysique.
Pour comprendre le caractère non empirique de cette approche, nous pouvons comparer ce paragraphe avec une autre version, celle de Denis Noble, le fameux physiologiste et biologiste des systèmes d’Oxford dans sa Musique de la Vie , publiée en 2006. Ce qui est un fait prouvé- les gènes « sont en vous et moi »- est maintenu ; l’interprétation a été réécrite, offrant un rôle radicalement différent au gène . Celui-ci n’est plus « actif » au sens fort du terme.
Les gènes sont piégés dans de grandes colonies, enfermés dans des êtres intelligents, modelés par le monde extérieur, communicant avec lui par des mécanismes complexes, grâce auxquels la fonction émerge, comme par magie de façon aveugle. Ils sont en vous en moi ; nous sommes le système qui leur permet la lecture de leur code ; et leur conservation est totalement dépendante de la joie que nous éprouvons en nous reproduisant. Nous sommes la raison ultime de leur existence.
Ces deux citations sont équivalentes d’un point de vue empirique, en ce qu’elles sont toutes les deux fondées sur l’observation de preuves expérimentales. Pourtant, elles expriment des choses totalement différentes concernant le rôle du gène. Alors, qui a raison ? Laquelle pourrait être qualifiée de « plus scientifique » ? Comment décider de ceci sur des bases scientifiques ? Comme Noble le fait remarquer : « personne ne peut faire d’expérience pour départager empiriquement ces deux citations.» Cette digression nous montre combien il est facile de faire intervenir des présuppositions métaphysiques dans ce qui devrait être un compte rendu scientifique des choses. Pour ceux qui en doutent encore, il peut être intéressant de réfléchir sur les paroles de William B. Provine, professeur de biologie à l’Université de Cornell. Que nous dit la biologie de l’évolution ? Ecoutons la réponse de Provine.
Laissez moi résumer ce que je pense de ce que la biologie de l’évolution nous dit haut et fort…Il n’y a pas de Dieu, pas d’objectifs, pas de force d’aucune sorte cherchant à atteindre un but. Il n’y a pas de vie après la mort. Lorsque je mourrai, je suis absolument certain que je serai mort. C’est la fin pour moi. Il n’y a pas de fondement ultime pour l’éthique, pas de sens ultime à la vie, et pas de libre arbitre pour les hommes non plus.
Je ne pense pas que ce soit de la biologie. Cela me paraît bien davantage être une projection d’une vision métaphysique du monde sur un pauvre support biologique matériel, qui est obligé d’écouter passivement et qui ne peut pas répondre.
Il est temps de conclure. Dans cette conférence, j’ai soutenu qu’il nous faut être très précautionneux dans le rejet prématuré de tout phénomènes téléologiques en biologie, qui sont souvent motivés par, soit des agendas métaphysiques masqués, ou bien par une mauvaise compréhension de ce qu’est vraiment la « téléologie ». Je suggère que le pendule a oscillé trop loin en excluant toute forme de téléologie en biologie. Alors que certains biologistes rejettent cette notion de façon axiomatique, par principe, j’aimerais argumenter que nous devrions mettre de tels dogmes métaphysiques de côté, et considérer les données empiriques. Ce que nous observons dans la nature, ce sont des schémas clairs. Qui ne correspondent directement à aucune notion métaphysique ou religieuse de téléologie- que ce soit aristotélicienne, kantienne, ou chrétienne. Pourtant, dans chaque cas, et tout particulièrement dans le cas de la doctrine chrétienne de la providence, ce qui est observé peut s’inscrire dans une vision plus large. Ceci ne prouve rien. Mais ceci suscite des questions intéressantes sur le regard que nous portons, sur la lecture que nous faisons du monde biologique.




