
Rappel
Rappelons quelques points exposés dans le dernier billet :
- L’apparition de nouveaux allèles est un événement unique qui a lieu dans un individu, mais il peut devenir commun dans une population par des processus variés, y compris par la dérive génétique et la sélection naturelle.
- Si de nouveaux allèles deviennent communs dans une population, ils peuvent changer les caractéristiques moyennes de cette population.
- Si l’échange d’allèles entre deux populations de la même espèce est bloqué ou réduit, alors les caractéristiques moyennes des deux populations peuvent diverger.
- Si le temps est assez long, ces processus peuvent amener à des différences assez significatives pour établir les deux groupes en deux espèces distinctes.
Ces points rappelés, nous pouvons maintenant considérer plus en détail les différentes manières dont un échange génétique entre des populations peut se réduire ou s’éliminer. Commençons par considérer le cas le plus simple : l’isolement géographique totale.
Les barrières géographiques
La séparation géographique de deux populations de la même espèce est une façon rapide et efficace d’arrêter l’échange d’allèles entre elles. Au moment de la séparation, les deux populations sont évidemment pleinement capables de se reproduire biologiquement, mais elles en sont empêchées par une séparation physique. Un exemple d’isolement géographique qui a amené à une spéciation que nous avons déjà vu, c’est celui des espèces variées de pinsons que Darwin a observées sur les îles Galapagos, au large des côtes de l’Amérique du Sud. La population de pinsons d’origine des Galapagos a été fondée par un groupe d’oiseaux qui est arrivé sur les îles du continent de l’Amérique du Sud, probablement à cause d’une tempête. Ces oiseaux, en tant que population, ont été coupés biologiquement de la population source du continent, puisque les Galapagos sont à des centaines de kilomètres au large. Une fois séparés, les deux groupes n’échangeaient plus d’allèles. Malgré leur appartenance à la même espèce, les deux populations sont désormais génétiquement différenciées, les différences de fréquences d’allèles s’étant accrues. Ce sont ces différences qui ont amené à un changement des caractéristiques moyennes de ce groupe à travers le temps, et finalement à la formation d’une nouvelle espèce.
L’effet fondateur
Dans de nombreux cas, ce processus d’accumulation des différences se déclenche dès le point de la séparation, à cause d’un phénomène que l’on appelle « l’effet fondateur ». Une petite population fondatrice est très souvent un échantillon non représentatif de la diversité génétique de la population source. Par exemple, considérons une population hypothétique composée de 36 individus. Chaque individu porte deux allèles d’un gène donné, et il y a quatre allèles différents de ce gène dans la population (représentés par les quatre couleurs) :

Remarquez que l’allèle jaune est le plus commun, suivi de l’allèle bleu. Les allèles rouge et violet, par comparaison, ne sont pas communs dans cette population. De fait, leur rareté signifie qu’il serait improbable que cette population ait un individu possédant deux allèles rouges à la génération suivante, par exemple. Afin d’avoir un tel individu, deux parents « porteurs » de l’allèle rouge devraient s’accoupler et transmettre l’allèle rouge à leur descendant. Ce n’est pas impossible, mais dans cette population ce serait improbable.
Supposons maintenant que quelques membres de cette population commencent une nouvelle population dans une île isolée. Seuls six individus commencent la nouvelle population, et les allèles qu’ils portent ne représentent par parfaitement les fréquences des allèles de la population source, plus large (des fréquences approximatives sont montrées pour la population source et la nouvelle population « fondatrice) :
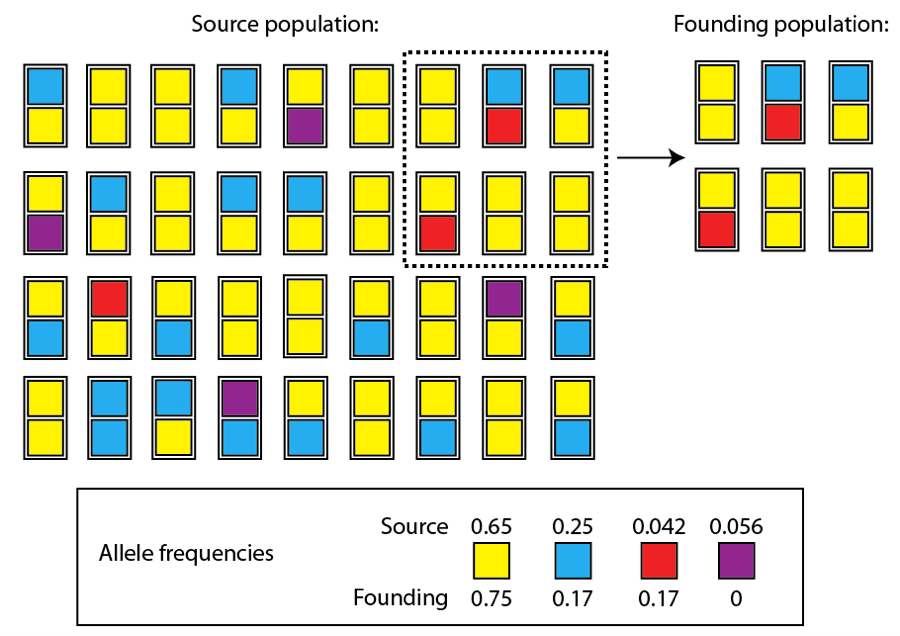
Nous pouvons voir en ce qui concerne les allèles communs, que la fréquence de l’allèle jaune a augmenté, et que celle de l’allèle bleu a baissé. Malgré ces différences, les allèles communs sont raisonnablement similaires en fréquence à ceux de la population source. Les allèles rares, cependant, ont vu leur fréquence changer plus significativement : l’allèle rouge est beaucoup plus commun dans la nouvelle population, tandis que l’allèle violet a complètement disparu.
Ces changements sont certes subtils, et les changements d’un gène ne seraient probablement pas suffisants pour précipiter un événement de spéciation entre les deux populations. Ces types de changements pourraient, cependant, être significatifs à long terme. Considérez l’allèle rouge dans la nouvelle population. Alors que cette population augmente en nombre, il sera bien plus probable que des individus portant deux allèles rouges apparaissent dans cette population que dans la population source. Si cette combinaison génétique a un avantage sélectif, alors la sélection naturelle pourra agir dans la nouvelle population. Cependant, dans la population source, cette combinaison génétique est bien plus improbable, prévenant ainsi la sélection naturelle d’agir sur cette combinaison d’allèles. A travers le temps, l’allèle rouge pourrait dominer la nouvelle population, mais rester rare dans la population source. De plus, il est probable que l’environnement sera différent d’une certaine manière pour ces deux populations, amenant à la sélection de combinaisons génétiques différentes. Ce qui pourrait être une combinaison avantageuse dans le continent peut ne pas l’être autant dans l’environnement insulaire, et vice versa. Une deuxième chose à prendre en compte, c’est que la nouvelle population, comme toute petite population, est bien plus sujette à la dérive génétique que la population source, plus large. La fréquence de l’allèle rouge peut augmenter dans la nouvelle population simplement par chance, et non par l’action de la sélection naturelle.
Pris ensemble, ces mécanismes peuvent mettre les deux populations sur des trajectoires différentes et à travers le temps, amener à des différences significatives entre elles. Si le temps est assez long, les différences peuvent être suffisamment importantes pour garder les populations séparées, même si elles devaient entrer en contact à nouveau. Dans ce cas, la plupart des biologistes classeraient les deux populations en deux espèces distinctes. Si cela est plus facile à faire pour des espèces qui ont été séparées pendant longtemps et ont accumulé des différences significatives (et qui ne se reproduisent plus entre elles, ou seulement rarement), il est notoirement difficile de le faire pour des populations qui se sont séparées récemment, et qui ne sont pas encore complètement isolées du point de vue de la reproduction. En tant que telle, ce qui constitue une « véritable espèce » plutôt qu’une « sous-espèce » ou « variété » est souvent le sujet d’une discussion ou d’un débat entre scientifiques. Et en effet, Darwin y a consacré beaucoup de temps dans ses travaux. Cette ambiguïté survient du mécanisme lent et graduel de divergence des espèces à partir d’une population ancestrale commune.
Pas seulement des différences
Etant donnée ce que nous venons de voir, vous pourriez avoir l’impression que la différence entre les espèces consiste le problème principal. Certainement, les différences sont vitales, puisque finalement ce sera l’accumulation de différences qui amènera à la formation de nouvelles espèces. Il est important de se rappeler, cependant, que pour des espèces proches, ces différences seront petites en nombre par comparaison avec les caractéristiques qui demeurent identiques dans les deux groupes. Au niveau génétique, nous pouvons l’illustrer en considérant un gène pour lequel il n’y a qu’un allèle dans la population source ; peut-être un allèle qui est passé par la sélection naturelle et qui a remplacé les’autres allèles. La nouvelle population n’héritera que de cet allèle, malgré la petite taille de l’échantillon que constitue le groupe, puisqu’il n’y a pas d’autres variantes dans la population. Le résultat est que la population insulaire sera identique à celle du continent dans ce trait, jusqu’à ce qu’un événement mutationnel (dans l’une ou l’autre population) autorise la possibilité d’un changement. Pour la plupart des traits, les mutations ne surviendront pas, puisque le mécanisme de copie de l’ADN est très précis. C’est ce qui gardera la plupart des traits communs aux deux populations. Le schéma attendu pour des espèces qui ont divergé récemment est donc des caractéristiques majoritairement identiques, avec quelques différences. Rappelons que c’est exactement ce schéma dans son contexte biogéographique qui a conduit Darwin à réfléchir à la possibilité de l’instabilité des espèces :
« Le fait le plus étonnant et le plus important pour nous en ce qui concerne les habitants de l’île, c’est leur affinité avec ceux du continent le plus proche, alors qu’ils ne sont pas de la même espèce. De nombreux exemples pourraient être donnés de ce fait. Je n’en donnerai qu’un, celui de l’archipel des Galapagos, situé sous l’Equateur, entre 800 et 1000km des côtes de l’Amérique du Sud. Ici, presque chaque produit de la terre et de l’eau porte l’empreinte évidente du continent américain. Il y a vingt-six espèces d’oiseaux terrestres, et parmi elles vingt-cinq sont recensées par M. Gould comme des espèces distinctes, dont on suppose qu’elles ont été créées ici. Pourtant l’affinité proche de la plupart de ces oiseaux aux espèces américaines dans tous leurs caractères, dans leurs habitudes, leurs comportement, les tons de leur voix, était manifeste. »
Notez que c’est le schéma composé d’ « affinités » évidentes (des caractères distinctifs communs) et de différences subtiles mais significatives que Darwin a observé. Les oiseaux en question étaient des espèces distinctes, mais ils conservaient « l’empreinte évidente » de leur héritage. Ce sont ces observations qui ont conduit Darwin à formuler l’hypothèse selon laquelle ces espèces de pinsons étaient le produit d’un événement de spéciation suite à l’isolement géographique.
Si l’isolement géographique est une situation directe qui peut amener à des barrières génétiques et à la formation de nouvelles espèces, la spéciation peut aussi arriver sans une séparation complète. Dans le prochain billet de cette série, nous examinerons un cas de spéciation qui n’a qu’une barrière géographique (et génétique) partielle ; un cas qui démontrera aussi le caractère flou de ce qui constitue exactement une espèce.
48 Articles pour la série :
- 01-L'évolution expliquée : Introduction
- 02-L'Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2) (Cet article)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l'homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l'humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour


