

« Mobius », sculpture mobile moderne de Julie Frith.
Note : Cette série de billets est une introduction de base pour les non spécialistes à la science de l’évolution. Vous pouvez voir l’introduction de cette série ici. Dans ce billet, nous introduisons la diversification cambrienne et voyons comment les « plans d’organisation » d’aujourd’hui permettent de prédire et de tester des patrons dans les données fossiles.
« L’explosion » cambrienne
Dans le dernier billet de cette série, nous avons vu certaines preuves en faveur de l’idée que deux anciens événements endosymbiotiques ont eu lieu (conduisant aux mitochondries et aux chloroplastes) et ont eu un impact profond sur l’évolution ultérieure de la diversité des eucaryotes. La diversification spectaculaire de groupes animaux pendant la période cambrienne, période qui s’étend de 542 à 488 millions d’années avant notre ère, est un second événement qui a profondément influé sur le futur de la vie animale sur terre.
La période cambrienne est particulièrement intrigante en ce qu’elle représente les premières données fossiles pour de nombreux groupes d’animaux qui ressemblent de façon perceptible à des animaux qui existent encore aujourd’hui. Par « perceptible », on n’entend pas bien sûr que ce que l’on voit de la période cambrienne est familier – la faune cambrienne est particulièrement différente des organismes d’aujourd’hui – mais plutôt que ce sont les premières apparitions dans les données fossiles de traits que l’on reconnaît comme étant caractéristiques de groupes d’organismes que l’on observe aujourd’hui. En d’autres termes, certains traits que nous observons chez les cambriens sont familiers, bien que la combinaison de traits que l’on observe chez les animaux Cambriens soit souvent différente de celle que l’on trouve chez les groupes modernes. Néanmoins, il y a un intérêt à déterminer quand les groupes que nous voyons aujourd’hui sont survenus – les premiers « arthropodes », ou les premiers « vertébrés », par exemple.
Si vous vous demandez si ceci introduit quelque chose comme une étude des données fossiles par un “biais contemporain”, vous avez raison : effectivement, les scientifiques utilisent les caractéristiques d’organismes contemporains pour tenter de regrouper par famille des organismes éteints. Avant de voir comme cela se manifeste dans notre étude des Cambriens, nous aurons à explorer quelques concepts sur les phylogénies.
Evolution et taxinomie
Les biologistes essaient de faire de la taxinomie, c’est-à-dire de regrouper des organismes dans des catégories logiques, depuis Carl de Linné, dans les années 1700. Etant donné le pouvoir explicatif de la théorie de l’évolution et sa place comme théorie fondatrice de la biologie actuelle, la taxinomie essaie désormais de regrouper les organismes selon leur parenté évolutive. Selon cette approche, les classifications les plus logiques sont dites monophylétiques, terme technique qui désigne simplement une population ancestrale commune ainsi que ses descendants. Un groupe monophylétique peut se reconnaître facilement si on l’imagine comme un mobile : un groupe monophylétique peut être coupé du mobile par un seul coup de ciseaux. Tout autre regroupement demanderait plusieurs coupes. Par exemple, pour la phylogénie suivante, le groupement A, B, C est monophylétique, mais pas le groupement B, C, D :
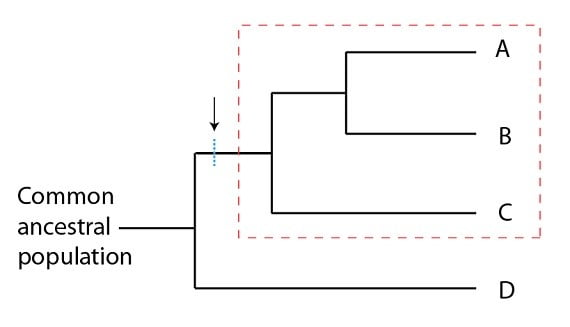
La ligne bleue coupe un groupe monophylétique d’une phylogénie. La science de classification des organismes (taxinomie) essaie d’organiser les espèces en groupes monophylétiques.
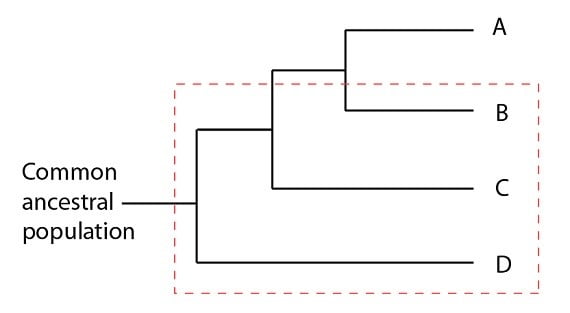
Le groupement B, C, D n’est pas monophylétique puisqu’il faudrait plusieurs “coupes” pour le retirer de sa phylogénie. Le groupement taxinomique correct qui inclut B, C, D doit aussi inclure A s’il se veut monophylétique.
Cependant, faire des groupes monophylétiques une base pour la taxinomie s’accompagne de défis. Un premier défi a lieu lorsque nous appliquons comme base cette tendance naturelle à utiliser des combinaisons de traits que l’on trouve dans des organismes d’aujourd’hui pour classer tous les organismes de l’histoire de l’évolution. Voyons un exemple pour comprendre quels sont les problèmes.
Que le premier “vrai” arthropode se signale !
Les arthropodes sont un groupe d’organismes très divers qui incluent des insectes contemporains, des crustacés, des arachnides (araignées, scorpions), entre autres. Les éléments que nous avons tendent à montrer que les arthropodes forment un groupe monophylétique. Tous les arthropodes vivants ont une suite de caractéristiques qui les définissent, comme un squelette externe et dur (exosquelette), des segments corporels spécialisés, des membres spécialisés. Si ces caractéristiques sont utiles pour la définition des arthropodes modernes, ces critères le sont moins lorsqu’on se tourne vers l’histoire évolutive des arthropodes. La raison en est simple : du point de vue de l’évolution, on ne peut pas s’attendre à ce que ces différents traits surviennent comme une unité, en une seule fois. On s’attendrait plutôt à ce que ces traits surviennent au cours du temps, dans le lignage qui conduit aux arthropodes d’aujourd’hui. Si c’est le cas, et si les populations s’éloignaient du lignage arthropode pour former des espèces alors que ces traits étaient acquis, on s’attendrait à trouver des espèces dans les données fossiles qui n’auraient pas l’apanage complet des caractéristiques des « arthropodes » mais seulement quelques unes :
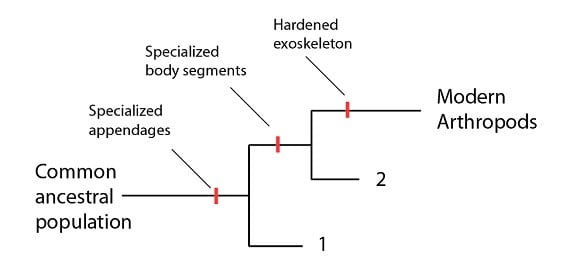
Par exemple, sur la base de la phylogénie ci-dessus, on peut s’attendre à trouver deux groupes d’organismes “type arthropode” dans les données fossiles : des espèces qui n’ont qu’un seul des trois traits (les membres spécialisés), ainsi qu’un second groupe qui aurait des membres spécialisés et des segments. Si de telles espèces (ou groupes d’espèces) existaient, cela nous informerait sur la façon dont l’ensemble des caractéristiques des arthropodes a été acquise au cours du temps, et brouillerait la distinction entre les arthropodes et les autres formes de vie. En effet, ces espèces représenteraient des « formes transitionnelles » au sens où elles ont un ensemble de caractéristiques intermédiaires qui indiquent les étapes que le lignage arthropode a traversées pour achever l’ensemble « moderne » de ses caractéristiques.
En d’autres termes, le groupe taxinomique des arthropodes est une classification arbitraire, puisque nous choisissons de couper un groupe monophylétique alors qu’il serait tout aussi approprié de le couper à un endroit différent, il y a plus longtemps, pour inclure plus d’espèces (ou plus tard, et en inclure moins).
Néanmoins, comprendre l’histoire évolutive d’un groupe monophylétique est très utile pour déterminer la chronologie de l’émergence des plans d’organisation des animaux, étape par étape. Dans notre prochain billet, nous explorerons cette idée en examinant certains animaux cambriens de façon plus détaillée.
Lectures complémentaires
Budd, G.E. (2008). The earliest fossil record of the animals and its significance Phil. Trans. R. Soc. B 363, doi: 10.1098/rstb.2007.2232
Budd, G.E. and Telford, M.J. (2009). The origin and evolution of arthropods. Nature 457, 812-817 doi:10.1038/nature07890
48 Articles pour la série :
- 01-L'évolution expliquée : Introduction
- 02-L'Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l'homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie. (Cet article)
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l'humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour



