
Article 30 sur un total de 48 pour la série :
![Photo credit: Davide Meloni [CC-BY-SA-2.0], via Wikimedia Commons)](https://scienceetfoi.com/wp-content/uploads/2015/01/1.jpg)
Photo credit: Davide Meloni [CC-BY-SA-2.0], via Wikimedia Commons)

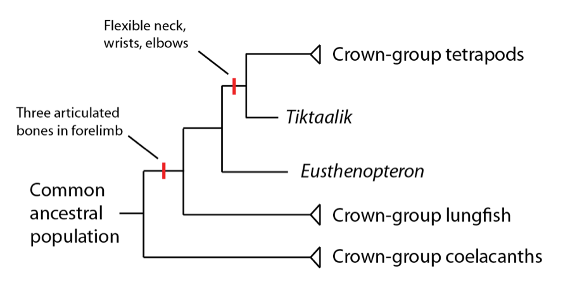
Phylogénie du groupe couronne des sarcoptégyriens : les tétrapodes d’aujourd’hui, les poissons pulmonés et les coelacanthes, leur dernière population ancestrale commune, et toutes les espèces descendantes extinctes. Les groupes souche des tétrapodes (marqués d’un « ? ») remplissent les « blancs » entre les groupes couronne du poisson pulmonaire et du tétrapode.
Les groupes souche des tétrapodes : du poisson à l’amphibien.
Il y a quelques années, ma famille a eu l’opportunité de visiter une exposition itinérante de fossiles dans un centre scientifique local. Je me rappelle très bien de notre journée, étant tiré d’un dinosaure spectaculaire à un autre par un enfant aux yeux grands ouverts qui les voyait en personne pour la première fois. Au milieu de ces géants impressionnants que mon fils admirait, cependant, se trouvait un étrange petit poisson qui a soudain éveillé mon excitation, au grand amusement de tous ceux qui m’entouraient à ce moment, qui l’ont à peine regardé. L’objet de mon exaltation était un membre du genre Eusthenopteron, un membre d’un groupe souche des tétrapodes proche de la dernière population ancestrale commune des poissons pulmonaires et des tétrapodes.
Comme nous l’avons vu dans les derniers billets, la clé qui permet de comprendre les transitions que fait un lignage au cours du temps est de chercher des groupes souche qui se séparent du lignage qui conduit au groupe couronne. Si des espèces de groupes souche ne sont probablement pas des ancêtres directs du groupe couronne, il est possible de trouver les parents qui sont plus proches du groupe couronne. Ces espèces, bien que n’étant pas les formes directes, ancestrales et transitionnelles qui conduisent au groupe couronne, préservent néanmoins les caractères transitionnels que le lignage du groupe couronne a transmis.
Peut-être que ceux qui autour de moi passaient à l’exposition de l’Eusthenopteron peuvent être excusés pour leur manque d’exaltation : après tout, un Eusthenopteron ne ressemble qu’à un petit poisson. Cependant, la structure de son squelette est tout sauf ennuyeuse pour un paléontologue. L’Eusthenopteron avait des os articulés dans ses nageoires avant – des os que nous reconnaissons chez les tétrapodes d’aujourd’hui comme l’humerus, le radius et l’ulna. Il s’agit des longs os qui font les membres avant dans le groupe couronne des tétrapodes ; mais chez l’Eusthenopteron, ces os sont courts et sont des soutiens pour les nageoires et non des membres (au sens de ceux qu’ont les tétrapodes). L’articulation de ces trois os dans le membre avant est un trait que l’Eusthenopteron partage avec les poissons pulmonaires mais pas avec les cœlacanthes, indiquant que cette caractéristique était présente dans leur dernière population ancestrale commune :

Les différentes espèces d’Eusthenopteron ont vécu il y a environ 385 millions d’années, au Dévonien, une période de l’histoire de la terre séminale pour les origines des tétrapodes, et une période fructueuse quant à la recherche d’espèces souches additionnelles pour les tétrapodes.
Tout est dans le poignet
Une fois que les chercheurs ont restreint un cadre temporel durant lequel un ensemble de transitions spécifiques ont eu lieu, il était possible de concentrer son effort pour y trouver de nombreuses espèces souches. Des tétrapodes (groupe souche) type amphibien comme les Acanthostega et les Ichthyostega apparaissent dans les données fossiles au Dévonien moyen, il y a environ 365 millions d’années. Ceci a permis de placer la transition des nageoires aux membres des tétrapodes entre des limites, du moment où le lignage tétrapode se sépare des Euthenopteron (il y a peut-être 400-390 millions d’années) jusqu’à l’apparition de ces tétrapodes amphibiens. De sorte que dans les dernières décennies, de nombreux efforts pour chercher des groupes souche de tétrapodes de cette période ont été menés pour éclairer la transition à l’état du groupe couronne tétrapode.
Une découverte qui a généré un intérêt significatif quand elle a été annoncée a été celle du Tiktaalik roseae, un groupe souche tétrapode d’abord découvert dans l’Arctique canadien en 2004. Tiktaalik est un fossile remarquable en ce qu’il est un vertébré à membre charnu avec des caractères type tétrapode particulièrement frappants, comme des coudes, des poignets flexibles (y compris des os de poignet qui correspondent à ceux du groupe couronne tétrapode), un cou flexible, et des membres avants (des nageoires avant ?) capables de soutenir son poids, ce qui a conduit à spéculer qu’il sortait peut-être de son habitat aquatique de temps en temps.

Tiktaalik roseae : un groupe souche dévonien aux caractères transitionnels entre les poissons à nageoire et le groupe couronne des tétrapodes (crédit : Wikimedia Commons)
D’autres découvertes ont largement rempli le vide entre les groupes du tétrapode et du poisson pulmonaire (voir par exemple la figure 6, dans Lu et al., 2012, un article qui décrit le tétrapode le plus ancien qui ait été trouvé jusqu’à maintenant, ou la figure 4 dans Swartz, 2012, un article qui décrit un tétrapode souche qui se détache du lignage du groupe couronne entre Eusthenopteron et Tiktaalik). Loin d’être un problème pour l’évolution, nous avons une excellente série de groupes souche tétrapodes qui révèlent les transitions pas à pas que le groupe couronne du lignage tétrapode a traversées, de la nageoire au membre.
Les groupes souche comme « formes transitionnelles ».
Bien que nous ayons répété et insisté sur l’idée que les espèces souche ne sont pas les ancêtres directs d’un groupe couronne, nous espérons qu’il est maintenant clair que les groupes souche sont en fait les « formes transitionnelles », formes dont il est raisonnable de penser que les données fossiles montreront, étant donné la nature rare du processus de fossilisation. Le fait qu’il soit hautement improbable qu’une espèce comme Tiktaalik soit l’ancêtre direct des tétrapodes modernes n’est en rien une objection qui empêcherait d’apprécier la combinaison de caractéristiques qu’elle possède, ni d’apprécier comme sa forme est (presque parfaitement) intermédiaire entre le vertébré à membre charnu et les tétrapodes. Objecter que les espèces souches ne sont pas des ancêtres directs, et donc ne nous informent pas sur les espèces d’aujourd’hui, c’est passer à côté du patron que les données fossiles nous présentent : des groupes d’espèces dans des ensembles qui s’incluent les uns dans les autres, mais des ensembles dont les frontières sont floues à chaque fois que nous avons suffisamment de données pour le voir.
Dans les prochains billets de cette série, nous suivrons le lignage tétrapode plus loin dans le futur, et examinerons la diversification des oiseaux et des mammifères.
48 Articles pour la série :
- 01-L’évolution expliquée : Introduction
- 02-L’Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l’homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l’humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour


