
Article 37 sur un total de 48 pour la série :

Dans le dernier billet de cette série, nous sommes arrivés au début des années 1950 dans l’histoire de la paléontologie hominienne, où Piltdown a été démasqué et l’interprétation correcte de l’Australopithecus africanus de Dart ainsi que du Pithecanthropus (Homo) erectus de Dubois, acceptée par la communauté scientifique. Les paléontologues savent désormais où chercher (en Afrique, comme Darwin l’avait suggéré), et ce qu’ils cherchent (des espèces ayant des corps de type humain et des capacités craniennes subhumaines). Lors des décennies suivantes, nombreuses découvertes seront faites, révélant que les hominiens constituent un large groupe composé de nombreuses espèces. Plutôt qu’une « échelle » d’espèces conduisant aux humains, la paléontologie hominienne a découvert un arbre touffu de parents proches.
Comme pour tous les groupes d’espèces fossilisées, déterminer leurs relations précises est un défi. Comme nous l’avons souligné auparavant dans cette série, la frontière entre les espèces est floue, la séparation se fait lentement et progressivement au cours du temps. Des espèces largement séparées sont faciles à identifier comme distinctes, mais à mesure que l’on recueille de plus en plus de données, les « fossés » se comblent, et faire des distinctions devient de plus en plus difficile. Dans une discipline telle que la paléontologie hominienne, ce problème est renforcé du fait de notre l’intérêt que nous portons à notre propre lignage ; ce qui signifie que nous avons mis en œuvre beaucoup de moyens pour découvrir autant de restes hominiens que possible. L’abondance de données rend difficile la résolution détaillée de l’image alors que les contours majeurs sont plus clairs. Cela conduit certains paléontologues à préfèrer délimiter un grand nombre d’espèces hominiennes fossiles, tandis que d’autres à regrouper les restes hominiens en un nombre plus restreint d’espèces. De façon informelle, ceux qui préfèrent un plus grand nombre d’espèces sont appelés les « splitters » (ceux qui séparent) et ceux qui préfèrent un nombre plus restreint d’espèces sont appelés les « lumpers » (ceux qui rassemblent). Le fait qu’un tel débat soit possible montre à quel point le panel des données qui sont disponibles sont robustes et s’échelonnent sans distinction claire.
La paléontologie hominienne : une petite esquisse des preuves actuelles
Globalement, en son état actuel, la paléontologie hominienne reconnaît plusieurs grands regroupements ou degrés avec, comme on s’y attend, des frontières floues. On trouvera une phylogénie consensuelle représentant la pensée actuelle ci-dessous, montrant la famille hominienne comme un arbre touffu, et mettant en valeur l’abondance de preuves que nous avons accumulées depuis l’époque de Dart et de Broom :
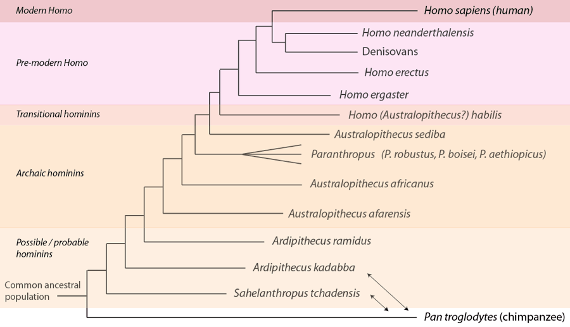
Phylogénie hominienne basée sur les preuves paléontologiques (et dans certains cas, génomiques) actuelles. Les séquences des génomes des humains, de Denisovan et de Neanderthal sont disponibles. Comme nous l’avons déjà vu, Sahelanthropus tchadensis et Ardipithecus kadabba ne sont peut-être pas des hominiens : ils pourraient être moins proches des humains que des chimpanzés, auquel cas leurs positions dans la phylogénie seraient à échanger avec celle des chimpanzés (indiqué par les flèches à double pointe). Plusieurs espèces hominiennes ne sont pas montrées. Voir le texte pour plus de détails.
Du passé au présent, le premier regroupement que nous rencontrons comprend des espèces que nous avons déjà vues : celles qui sont proches de la dernière population ancestrale commune aux humains et aux chimpanzés. Ce groupe, que l’on appelle les « hominiens possibles et probables » comprend les espèces « possiblement » hominiennes Sahelanthropus tchadensis, Orrorin tugenensis et Ardipithecus kadabba, et seulement une espèce « probablement » hominienne, Ardipithecus ramidus. Comme nous l’avons déjà noté, les indices en faveur de la bipédie sont plus forts en ce qui concerne Ardipithecus ramidus. Si les « splitters » considèrent que ce sont deux espèces distinctes, les « lumpers » les rassemblent sous le nom d’Ardipithecus ramidus sensu lato, « sensu lato » signifiant « au sens large ». En général, ce regroupement a un petit volume crânien (moins de 350 cm3) et des signes de bipédie.
Le second regroupement, « les hominiens archaïques », comprend les Australopithèques comme l’Australopithecus africanus de Dart et une espèce plus ancienne, Australopithecus afarensis, dont l’exemple le plus connu a été surnommé « Lucy ». Il comprend aussi les hominiens archaïques dits robustes, les différentes espèces assignées à Paranthropus (comme le Paranthropus robustus de Bloom). En général, on voit dans ce regroupement plus d’indices en faveur d’une bipédie occasionnelle (c’est-à-dire facultative) et une augmentation du volume crânien pouvant aller jusqu’à 500cm3. Si les « splitters » voient de nombreuses espèces dans ce regroupement, les « lumpers » n’y voient que A. afarensis, A. africanus, et une espèce comprenant tous les Paranthropus.
Le troisième regroupement, les “hominiens de transition”, contient les membres Homo les plus anciens que l’on connaisse, comme Homo habilis. Certains scientifiques cependant, remettant en question leur nature transitionnelle, ne placent pas cette espèce parmi les Homo mais la décrivent plutôt comme un australopithèque, Australopithecus habilis. Le volume crânien des habilis se situe entre 500cm3 et environ 700cm3. Homo rudolfensis, une espèce similaire, est reconnue par les « splitters », mais regroupée sous le terme de H. habilis sensu lato par les « lumpers ». Ces espèces (ou cette espèce) révèlent des signes croissants de la bipédie, mais pas une pratique complète d’un mode de vie bipède (c’est-à-dire d’une bipédie obligatoire), comme chez les Homo plus récents. Ces groupes utilisaient aussi des outils en pierre oldowayens, une technologie d’outils en pierre relativement simple ; cette technologie tire son nom du site où elle a été découverte pour la première fois.
Le quatrième regroupement, “Homo pré-moderne” comprend Homo ergaster et l’Homo erectus de Dubois, l’espèce Homo la plus ancienne qui soit complètement bipède. Si les « splitters » reconnaissent les ergaster et erectus comme deux espèces séparées, les « lumpers » les placent ensemble comme Homo erectus sensu lato. Ce regroupement comprend également les espèces plus familières Néanderthal et Denisovan, dont nous possédons les séquences complètes de génome. Leur volume crânien commence à 700cm3 pour les anciens Homo ergaster/erectus, et peut atteindre celui des humains modernes (environ 1400cm3) (et dépasse le volume moyen de l’humain moderne dans le lignage néanderthal, où l’on trouve des crânes d’un volume de 1600cm3). Assez tôt à cette étape, nous voyons se développer une technologie de l’outil en pierre acheuléenne plus complexe.
Le dernier regroupement, “Homo moderne”, ne contient que notre propre espèce, Homo sapiens, qui entre dans les données fossiles il y a environ 200 000 ans. Les « splitters » ont cependant soutenu qu’il y avait une sous-espèce, Homo sapiens idaltu, il y a environ 160 000 ans.
Des ancêtres, peut-être : des parents, certainement
Cette diversité des fossiles complique l’histoire détaillée de notre évolution, en ce sens qu’on ne peut pas être absolument certain lequel (s’il en est un) de ces groupes sont des ancêtres directs de notre propre espèce, et en même temps en clarifie le paysage global, puisqu’ils sont nos proches parents, même s’ils ne sont pas nos ancêtres directs. Globalement, on peut penser que notre lignage a traversé ces différentes étapes tour à tour : d’un ardipithèque (ou un parent proche) à un australopithèque (ou un parent proche), et ainsi de suite en passant par les hominiens de transition puis par l’Homo pré-moderne et enfin moderne. Sur le chemin, le volume crânien de notre lignage s’est développé en moyenne, notre lignage a acquis l’utilisation d’outils plus sophistiqués et, il y a environ 200 000 ans, a pris notre forme actuelle en Afrique, avant notre expansion sur le globe.
Dans le prochain billet de cette série, nous continuerons l’histoire de notre espèce qui s’étend hors d’Afrique et qui va rencontrer les descendants de lignages parents qui avaient quitté l’Afrique avant nous.
Pour en savoir plus
- Wood, B. and Lonergan, N. (2008). “The hominin fossil record: taxa, grades and clades.” Journal of Anatomy 212: 354 – 376.
- James Kidder: “The Human Fossil Record.”
48 Articles pour la série :
- 01-L’évolution expliquée : Introduction
- 02-L’Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l’homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l’humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour




