
Article 22 sur un total de 48 pour la série :
Dans le dernier billet de cette série, nous avons introduit le concept d’homoplasie : une similitude de forme dans deux lignages qui survient par des événements indépendants. Nous avons vu la dernière fois l’exemple du vol actif des oiseaux et des chauves-souris obtenu de façon indépendante par une convergence évolutive, les chauves-souris ayant des ailes faites de membranes et les oiseaux, de plumes. Puisque la dernière population ancestrale commune des oiseaux et des chauves-souris n’avait pas de vol actif, c’est un bon exemple d’homoplasie survenue par une convergence évolutive.
Sous cet événement de convergence, cependant, il y a une connexion plus profonde. Les chauves-souris et les oiseaux sont des tétrapodes, c’est-à-dire des organismes avec quatre membres et une colonne vertébrale. La structure corporelle du tétrapode était déjà une caractéristique de leur dernière population ancestrale commune, et a été maintenue dans les deux lignages. En tant que telles, lorsqu’on les considère strictement comme des membres antérieurs, les ailes des chauves-souris et celles des oiseaux sont des structures homologues. Chez les oiseaux comme chez les chauves-souris, les membres antérieurs ont été formés par la sélection naturelle pour le vol de différentes façons, mais le point de départ pour les deux était une structure homologue. En d’autres termes, sous l’événement de convergence du vol actif des chauves-souris et des oiseaux se trouve une homologie plus profonde : le membre sur lequel les deux lignages ont indépendamment construit une aile. Pour le représenter sur une phylogénie, nous placerions la structure corporelle du tétrapode avant la divergence de tous les tétrapodes, et ferions du vol actif deux événements différents sur les lignages appropriés :
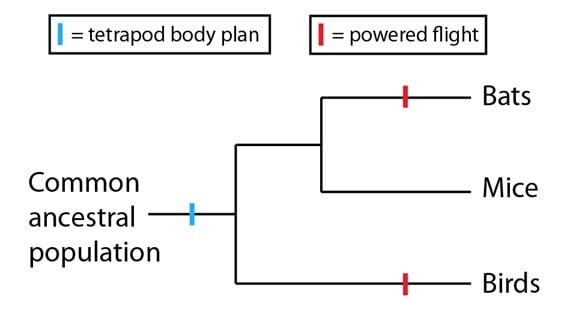
Des événements de convergence sous-tendus par des homologies plus profondes forment un patron qui est vu de temps à autres dans l’évolution. En fait, ces homologies plus profondes augmentent les chances que des événements de convergence arrivent, puisqu’elles donnent une base commune que des lignages séparés peuvent utiliser pour des innovations indépendantes. Pour les chauves-souris et les oiseaux, les adaptations qui ont conduit au vol étaient possibles parce que les deux lignages avaient des membres antérieurs qui pouvaient être modifiés au cours du temps d’une fonction à une autre. Si cet exemple est au niveau anatomique, ces espèces de « prédispositions » et les événements de convergence qui en surviennent peuvent également être observés au niveau moléculaire.
Les yeux.
Comme nous l’avons mentionné dans le dernier billet de cette série, les yeux camérulaires sont l’un des exemples les plus frappants de l’évolution convergente, puisqu’ils sont apparus indépendamment dans plusieurs lignages (les exemples les plus courants sont les vertébrés, les céphalopodes comme les pieuvres et les calamars, et certaines méduses). Les yeux camérulaires ont une couche cellulaire sensible à la lumière (la rétine) ainsi qu’une lentille qui concentre la lumière sur la rétine. Expliquer la distribution des yeux camérulaires chez ces trois groupes implique d’invoquer trois événements de convergence sur leur phylogénie (les « cnidaires » sont le groupe dans lequel on trouve les méduses) :
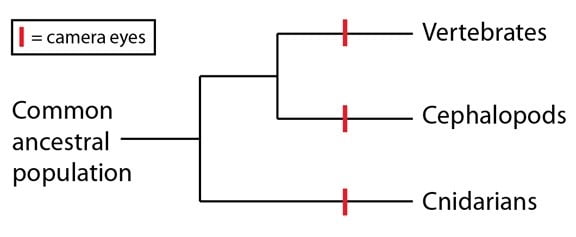
A première vue, il semble absolument improbable que trois lignages apparentés de façon très distante convergent indépendamment à une structure aussi remarquable que celle des yeux camérulaires. Il se trouve, cependant, qu’une homologie clé entre les trois groupes en a augmenté les probabilités : les molécules qui jouent le rôle de détecteurs de lumière.
Dans sa forme la plus basique, la sensation de l’environnement extérieur requiert de l’environnement qu’il induise un changement dans les cellules. Ainsi, la sensation de la lumière requiert un changement induit par la lumière d’un certain type. Les molécules clés de cette fonction dans les trois groupes ci-dessus sont des protéines que l’on appelle des opsines et leurs compagnons chimiques, les rétinals .Chaque protéine opsine a un rétinal, qui lui est lié, et ils jouent ensemble le rôle d’un détecteur de lumière. Les rétinals changent de forme lorsqu’ils interagissent avec la lumière (c’est-à-dire lorsqu’ils absorbent un photon, représenté par un gamma dans le diagramme ci-dessous). Ce changement altère à son tour la forme de l’opsine attachée au rétinal :
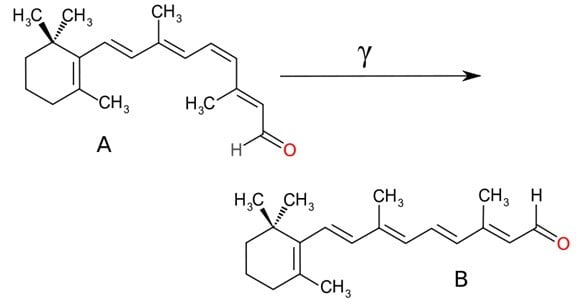
Source: http://en.wikipedia.org/wiki/File:RetinalCisandTrans.svg
Le changement de forme de l’opsine affecte le flux de la charge électrique dans les cellules responsables de la sensation de la lumière, et ces changements de la charge électrique sont ce que le cerveau perçoit et interprète comme la « lumière ».
Le système opsine/rétinal de détection de lumière est très répandu ; en fait, tous les animaux qui peuvent détecter la lumière utilisent ces molécules comme base physique, qu’ils aient des yeux camérulaires ou d’autres types d’yeux (les yeux composés par exemple, ou de simples ensembles de cellules sensibles à la lumière). C’est une preuve solide que le système opsine/rétinal précède la divergence des trois groupes que nous considérons :
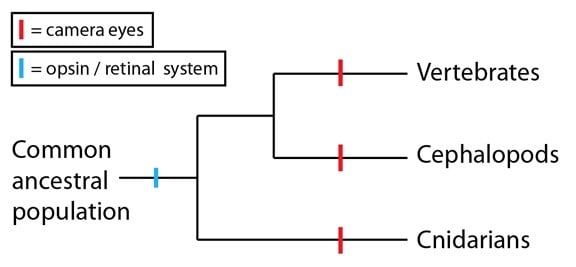
Une fois que nous savons cela, nous voyons que le développement des yeux camérulaires dans ces lignages n’est pas si improbable qu’on l’aurait pensé au départ. Dans les trois cas, ces lignages ont construit un œil camérulaire autour d’un système moléculaire détecteur de lumière préexistant. Les yeux camérulaires en tant que tels sont convergents, mais ils sont basés sur une homologie sous-jacente plus profonde qui a augmenté les probabilités de leur apparition par des modifications successives d’un système ancestral. Et comme nous l’avons vu pour les ailes d’un oiseau et d’une chauve-souris, il y a des différences entre les yeux camérulaires de ces lignages qui soutiennent l’hypothèse que ce sont les résultats d’événements de convergence (l’exemple le plus connu étant le nerf des yeux des vertébrés et celui des céphalopodes qui ont une orientation différente).
L’entendre c’est le croire.
On peut voir un second exemple de la “prédisposition moléculaire” conduisant à la convergence dans la mécanique moléculaire qui sous-tend une forme de perception sensorielle différente : celle des ultrasons par l’audition, requise pour l’écholocalisation chez les chauves-souris et les cétacés à dents. Les deux groupes utilisent une écholocalisation à haute fréquence pour naviguer et chercher une proie dans un environnement où la perception visuelle est limitée voire impossible. Il y a de fortes preuves qui montrent que le développement de l’écholocalisation chez ces deux groupes de mammifères très divergents est dû à l’évolution convergente ; il n’y a pas d’autre mammifère apparenté de plus près à l’un de ces deux groupes qui ait une telle capacité.
La base cellulaire/moléculaire qui détecte les sons chez les mammifères est un ensemble de cellules dans l’oreille prolongées par des extensions qui ressemblent à des cheveux (que l’on appelle cils cellulaires), qui vibrent en réponse à différentes longueurs d’onde de son. Les cils cellulaires changent aussi de longueur et de propriétés vibratoires en réponse à des stimuli auditifs différents. Les vibrations sont utilisées pour changer le flux de la charge électrique de ces cellules, conduisant finalement des signaux au système nerveux, que le cerveau perçoit comme des sons. Tous les mammifères utilisent une protéine que l’on appelle prestine dans leur système auditif. La prestine est une « protéine moteur » qui peut changer la forme d’une cellule en déplaçant les structures internes ; et les mammifères l’utilisent pour modifier les cils cellulaires en réponse au son.
On sait que le système cil cellulaire/prestine précède tous les mammifères, donc il n’est pas surprenant que les cétacés à dents et les chauves-souris utilisent ce système pour entendre. Ce qui est intéressant, cependant, c’est que la prestine ait été formée dans ces deux groupes de manière indépendante par la sélection naturelle pour entendre les sons à haute fréquence (les ultrasons), plus utile pour l’écholocalisation. De fait, sur une phylogénie restreinte aux séquences de prestine, celles des chauves-souris et celles des cétacés à dent semblent être les plus proches, ce qui est en forte contradiction avec l’arbre généalogique des espèces des chauves-souris et des cétacés. Un examen plus approfondi, cependant, montre que ces frappantes similitudes sont le résultat d’une convergence évolutive, et non d’un ancêtre commun plus récent. Dans les deux cas, la protéine de prestine pouvait s’accorder aux longueurs d’ondes ultrasoniques, et des événements mutationnels similaires (mais non identiques) ont été sélectionnés dans les deux lignages en cours de route, ce qui est un exemple de plus d’une « profonde homologie » favorisant des événements de convergence indépendants.
En résumé : l’évolution comme processus non hasardeux.
Une mauvaise conception de l’évolution que je rencontre souvent, c’est qu’elle est surtout un processus hasardeux, principalement influencé par des événements aléatoires. Si nous avons déjà montré que l’évolution a un élément non-hasardeux fort (la sélection naturelle), notre propos sur la convergence évolutive démontre un peu plus que l’évolution peut se répéter de façon importante. Lorsque la sélection naturelle affecte des groupes apparentés de loin d’une façon similaire, nous observons souvent des résultats similaires. Ceux-ci sont dans de nombreux cas favorisés par une histoire (homologie) et atteints par des voies similaires mais non identiques (ce qui démontre que la contingence et la chance sont également présentes). L’évolution est ainsi un équilibre entre événements contingents (des mutations et d’autres événements dus à la chance), et événements non-contingents (la sélection, la convergence évolutive).
Dans le prochain billet de cette série, nous reviendrons à l’écholocalisation des chauves-souris pour explorer la façon dont l’évolution d’une espèce peut être contrainte par celle d’une autre espèce qui a une relation proche avec elle, un phénomène que l’on nomme la coévolution.
48 Articles pour la série :
- 01-L’évolution expliquée : Introduction
- 02-L’Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l’homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l’humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour

