
Article 27 sur un total de 48 pour la série :
Note : Cette série de billets se veut être une introduction de base à la science de l’évolution pour les non spécialistes. Vous trouverez l’introduction de cette série ici. Nous introduirons dans ce billet les concepts de « groupes couronne » et de « groupes souche », et les appliquerons à la faune cambrienne pour révéler les étapes du lignage arthropode qui ont conduit au plan d’organisation arthropode moderne.
Dans notre dernier billet, nous avons introduit l’idée que ce que nous définissons aujourd’hui comme un “plan d’organisation” ou “une suite de caractéristiques partagées par une famille monophylétique” présente en fait des problèmes pour la classification des organismes passés. La raison, nous l’avons vu, en est que toute suite de caractéristiques modernes ne peut être survenue en une seule fois ; on ne peut s’attendre qu’à les voir survenir par étapes, pas à pas.
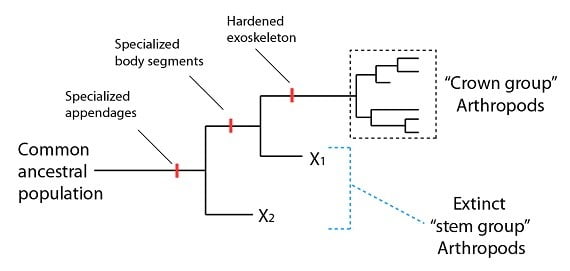
Revenons à notre discussion sur les arthropodes et voyons comment les biologistes gèrent ce problème lorsqu’ils se confrontent aux données réelles. En voici l’approche générale : il s’agit de commencer par les caractéristiques d’une famille d’espèces monophylétiques qui a au moins deux espèces représentées aujourd’hui. Ces espèces, leur ancêtre commun et tous les descendants de cet ancêtre commun forment ce qu’on appelle un « groupe couronne » sur une phylogénie (on l’abrège souvent par un triangle pour représenter de nombreuses espèces). Ainsi, nous pouvons définir le groupe couronne des arthropodes comme le groupe qui contient le dernier ancêtre commun de toutes les espèces arthropodes qui vivent encore et ses descendants (en vie ou pas). D’autres familles éteintes qui ont certaines caractéristiques du groupe couronne (« crown group ») sont alors classifiées comme des espèces appartenant au « groupe souche » (« stem group ») :

Les onychophores font partie de la famille des arthropodes qui vivent dans des lieux tropicaux et des régions tempérées de l’hémisphère sud. Bien que segmentés, il leur manque certains membres spécialisés et certaines caractéristiques segmentées des arthropodes du groupe couronne.
Les onychophores font partie de la famille des arthropodes qui vivent dans des lieux tropicaux et des régions tempérées de l’hémisphère sud. Bien que segmentés, il leur manque certains membres spécialisés et certaines caractéristiques segmentées des arthropodes du groupe couronne.
Une question se pose alors : un « groupe souche » arthropode est-il vraiment arthropode ? Oui et non – de telles espèces n’appartiennent pas au groupe couronne arthropode, puisqu’elles ne possèdent pas toutes les caractéristiques qui définissent les arthropodes d’aujourd’hui (comme l’ancêtre commun et ses descendants). Cependant elles sont plus proches du groupe couronne des arthropodes que de tout autre groupe monophylétique qui ait des représentants en vie, et possèdent au moins quelques caractéristiques du groupe couronne. En tant que telles, elles sont des arthropodes en un sens, mais on les décrit mieux en les nommant groupe souche.
Elargissons cet exemple pour y inclure un autre groupe couronne : celui des onychophores (Onychophora). Les onychophores partagent certains caractères communs avec les arthropodes (comme les membres et les segments) ; ils manquent certains autres (par exemple la spécialisation complexe des segments).
Les analyses phylogénétiques (qui utilisent la morphologie comparative, les données fossiles, et les séquences ADN) rapprochent constamment les onychophores des arthropodes. Puisque les onychophores sont des espèces en vie, on peut définir un groupe couronne pour eux également (comme auparavant, on y inclura le dernier ancêtre commun de toutes les espèces en vie des onychophores ainsi que toutes les espèces qui en descendent). Comme pour les arthropodes, on voit lorsqu’on examine les données fossiles des espèces éteintes qui ont certaines caractéristiques qui définissent le groupe couronne des onychophores. On peut donc placer ces espèces sur la branche du lignage des onychophores :

Il ressort de cette phylogénie que la distinction entre un groupe souche arthropode et un groupe souche onychophore deviendra de plus en plus floue au fur et à mesure que les deux lignages convergent (en remontant d’aujourd’hui jusqu’à leur ancêtre commun). Les espèces que l’on trouve dans les données fossiles peuvent être assignées comme souche à l’un ou l’autre des lignages en se basant sur leur suite de caractéristiques, en voyant si elles semblent plus proches du lignage arthropode ou du lignage des onychophores.
Les groupes souche arthropodes dans la période cambrienne
Une fois cela compris, nous pouvons nous tourner vers les espèces que nous observons dans les données fossiles cambriennes, exemples d’un groupe souche arthropode. Le grand prédateur cambrien Anomalocaris (littéralement, crevette anormale) est bien connu. Anomalocaris possède un certain nombre de caractères qu’il partage clairement avec le groupe couronne arthropode, comme la combinaison de grands yeux composés, des membres joints spécialisés, une segmentation spécialisée, et d’autres encore. Cependant, il lui manque un exosquelette dur (seuls ses membres sont durs). Cette suite de caractéristiques le rapproche du groupe couronne arthropode (c’est-à-dire que l’on placerait Anomalocaris sur le X de la phylogénie ci-dessus), et nous informe sur l’état des caractéristiques présentes à l’époque où le lignage qui a conduit à l’Anomalocaris s’est séparé du lignage qui a conduit au groupe couronne arthropode (c’est-à-dire qu’un exosquelette complètement dur est l’une des dernières caractéristiques acquise par la population ancestrale conduisant au groupe couronne arthropode).
D’autres espèces cambriennes montrent encore moins de caractéristiques communes au groupe couronne arthropode, et manifestent pourtant au moins quelques caractères en commun. L’étrange groupe d’espèces que l’on appelle Hallucigenia en est un exemple. Les Hallucigenia partagent les membres spécialisés ainsi que la segmentation spécialisée avec le groupe couronne arthropode ; mais d’autres caractères comme l’exosquelette dur (que ce soit un membre ou le corps tout entier) sont absents. En d’autres termes, ces espèces sont des souches arthropodes qui ressemblent plus à ce qu’on attendrait des souches onychophorans.
La mise en place des plans d’organisation animaux, pas à pas
On peut résumer ces découvertes comme suit :
Ce que nous observons comme l’émergence d’une nouvelle unité taxinomique (« phylum », « famille », « genus », etc) est en quelque sorte arbitraire (puisqu’elle décrit un continuum) et est n’est décidée qu’en prenant du recul, en se basant sur les caractéristiques de familles monophylétiques d’aujourd’hui.
Pour le meilleur et pour le pire, la taxinomie essaie de ranger les anciennes espèces dans des catégories modernes. Le fait que les anciennes espèces brouillent les distinctions que l’on fait aujourd’hui entre des groupes taxinomiques d’aujourd’hui (comme les arthropodes et les onychophores) montre que ce que nous reconnaissons comme de larges groupes taxinomiques (comme ce que nous appelons « phyla ») sont en fait mieux décrits comme des groupes monophylétiques dans des imbrications d’ensembles.
Les données fossiles soutiennent l’idée d’une acquisition qui s’est fait petit à petit des caractéristiques que nous voyons dans les groupes d’aujourd’hui et que nous utilisons par conséquent pour la classification.
Les caractères clés des arthropodes (segmentation spécialisée, membres spécialisés, exosquelettes durs, yeux composés, ainsi que d’autres encore) n’apparaissent pas d’un coup dans quelques espèces que l’on trouve dans les données fossiles. Dire que les arthropodes surviennent et se diversifient pendant la période cambrienne n’est pas dire que les arthropodes arrivés à pleine maturité (c’est-à-dire ceux qui appartiennent au groupe couronne) sont soudainement apparus, venant de nulle part. Nous voyons plutôt une série d’espèces qui démontre que les caractéristiques propres au groupe « couronne » arthropode ont été acquises sur le long terme. Si des espèces du groupe souche ne sont probablement pas des ancêtres directs de l’espèce du groupe couronne, leur présence dans les données fossiles (et la hiérarchie imbriquée que leurs caractéristiques produisent) nous donne les moyens de déterminer l’ordre dans lequel les caractéristiques propres ont été assemblées.
Pour en savoir plus
Budd, G.E. (2008). The earliest fossil record of the animals and its significance Phil. Trans. R. Soc. B 363, doi: 10.1098/rstb.2007.2232
Budd, G.E. and Telford, M.J. (2009). The origin and evolution of arthropods. Nature 457, 812-817 doi:10.1038/nature07890
48 Articles pour la série :
- 01-L’évolution expliquée : Introduction
- 02-L’Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l’homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l’humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour


