
Article 14 sur un total de 48 pour la série :
Rappelons ce que nous avons vu dans les derniers billets de cette série : la spéciation commence quand une barrière au flux d’allèles surgit et sépare une population en deux. Une fois que les deux populations ne peuvent plus échanger d’allèles (ou ne le font qu’à un taux réduit), des différences s’accumulent dans les deux populations alors qu’elles ne communiquent plus entre elles. De plus, nous avons vu que l’effet de fondation peut lancer ce processus, puisqu’il peut conduire à des différences lors de la séparation. Une fois que deux populations sont séparées et ont cessé de se reproduire entre elles, les nouveaux allèles qui surviennent dans une population ne seront pas étendus à l’autre, ce qui amène à des différences croissantes avec le temps.
Ce qu’il nous faut souligner ici, c’est que lorsque la spéciation commence, elle part d’une population ancestrale commune. Cela signifie que deux populations, au début, ont des génomes (presque) identiques. En effet, seuls les quelques gènes qui ont des fréquences d’allèles différentes dues à l’échantillonnage imparfait lors de la séparation des populations seront différents au début. A partir de ce point de départ, les deux populations commenceront à progressivement accumuler des différences – mais ces différences seront minuscules. Le patron général sera de façon prédominante celui de séquences identiques, contenant un petit nombre de différences.
Une analogie : les génomes sont comme des textes anciens.
Peut-être qu’une analogie pourrait aider. Avant l’invention de l’imprimerie, les manuscrits du monde ancien étaient copiés par des scribes. Bien qu’un bon scribe fût capable de donner une copie presque exacte, de petites erreurs de copie étaient inévitables. Ces changements, cependant, n’étaient pas assez significatifs pour empêcher la reconnaissance des copies – la vaste majorité du texte était correcte. Une fois qu’une copie était faite (contenant les petites erreurs), elle servait souvent de modèle à d’autres copies. Dans ce cas, les erreurs étaient copiées, puisque que le prochain scribe essayait aussi de copier le manuscrit aussi fidèlement que possible (même si à son tour, il pouvait introduire de nouvelles erreurs).
Le génome peut être pensé comme un “texte” transmis par des copies et, possiblement, avec des erreurs de copie. Cependant, comme pour toute analogie, il y a d’importantes différences. Si les scribes humains interagissent avec la signification du texte qu’ils copient, ce n’est pas le cas des « scribes » du génome – des enzymes qui copient les séquences d’ADN fondées sur l’appariement de monomères. Cela signifie que si des erreurs faites par des scribes humains tendent à préserver une certaine signification (même s’il s’agit d’une signification altérée), les enzymes qui répliquent l’ADN ne vérifient pas si la signification (c’est-à-dire la fonction) est préservée pendant la copie. (La vérification fonctionnelle d’une séquence d’ADN viendra plus tard, alors que cet organisme particulier se développe (ou pas) et se reproduit (ou pas). En d’autres termes, la sélection naturelle est la vérification de la « signification » d’une séquence d’ADN.
Pour continuer l’analogie, nous pourrions considérer le génome d’un organisme comme un livre, avec des chapitres, des paragraphes et des phrases. Pour un génome, les « chapitres » seraient les séquences de chromosomes ; les paragraphes seraient les gènes ; et les phrases seraient les sous-éléments des gènes. Nous pourrions aussi considérer les éditions d’un livre comme un événement de réplication. Par exemple, considérons deux éditions indépendantes du même manuscrit. Elles seraient, bien sûr, presque identiques – mais supposons que les deux éditions aient des fautes de frappe spécifiques qui n’altèrent pas beaucoup la signification du texte, et passant ainsi inaperçues des éditeurs : dans la première édition à la page 14, et dans la seconde, à la page 23 :
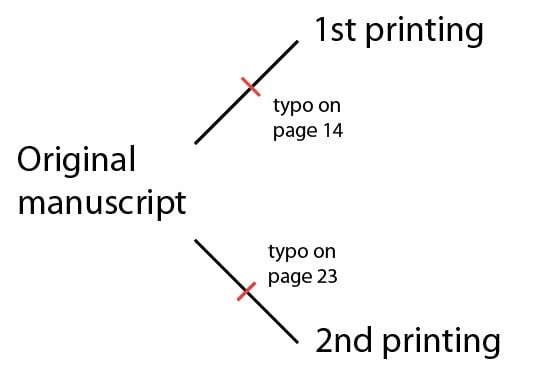
Supposons maintenant que le manuscrit original est perdu, et que la troisième édition est composée d’après une copie de la première édition. Cette nouvelle édition aurait la faute de frappe à la page 14, et n’importe quelle nouvelle faute de frappe (par exemple, à la page 8) :
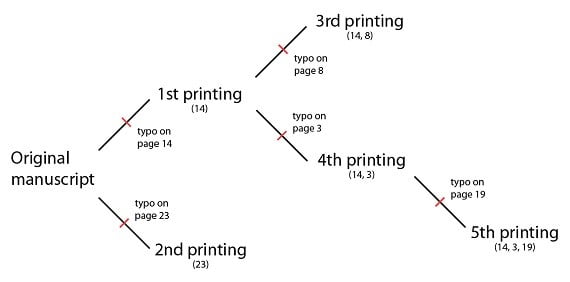
Imaginons maintenant qu’une sixième édition, auparavant inconnue, est découverte. Cette édition possède la même faute de frappe sur les pages 14 et 3 que l’on trouve dans la quatrième édition, ainsi qu’une faute de frappe unique, que l’on n’a pas encore vue dans les autres éditions, sur la page 5.
Il existe plusieurs explications possibles de la provenance de la sixième édition, l’une d’entre elles étant plus probable que les autres. Dans l’ordre croissant de probabilité, certaines options sont :
- La “sixième édition” est en fait un livre écrit par un autre auteur qui n’est pas la copie du manuscrit original duquel les cinq éditions sont des copies.
- La sixième édition est une copie directe du manuscrit original, mais l’éditeur a fait indépendamment des autres manuscrits les mêmes erreurs, en y ajoutant l’erreur de la page 5.
- La sixième édition est une copie directe de la première édition, mais l’éditeur a fait indépendamment des autres manuscrits les mêmes erreurs ainsi qu’une nouvelle erreur sur la page 5.
- La sixième édition est une copie directe de la quatrième édition, mais l’éditeur a fait une nouvelle erreur à la page 5 en copiant.
Il va de soi que la première option ne serait pas considérée sérieusement par les spécialistes, étant donné le texte quasiment identique partagé entre l’édition découverte et les autres éditions connues. La deuxième option requiert que deux événements rares (des fautes de frappe) soient arrivés indépendamment l’un de l’autre, deux fois, dans deux éditions. En tant que telle, elle est moins probable que la troisième option, qui ne requiert qu’un seul événement rare, qui serait arrivé deux fois indépendamment l’un de l’autre. La quatrième option est bien sûr la meilleure, parce qu’elle ne requiert pas que deux événements rares arrivent deux fois dans des copies indépendantes. Dans ce scénario, ce qui explique les caractéristiques de la sixième édition c’est qu’elle est une copie (imparfaite) de la quatrième édition :

En termes scientifiques, cette option est la plus probable, ou la plus parcimonieuse : elle offre une explication de la provenance de la sixième édition avec le nombre le plus faible d’événements.
Retour à la biologie
Considérons maintenant les événements de la spéciation à la lumière de notre analogie. Lors de la séparation, les deux populations ont des « livres » quasiment identiques (les génomes). Alors que les lignages font leur chemin séparément, les « fautes de frappe » (les mutations) peuvent arriver dans des gènes ensuite transmis aux descendants de ce lignage, comme nous l’avons vu dans le cas des fautes de frappe qui s’accumulaient dans les textes copiés :
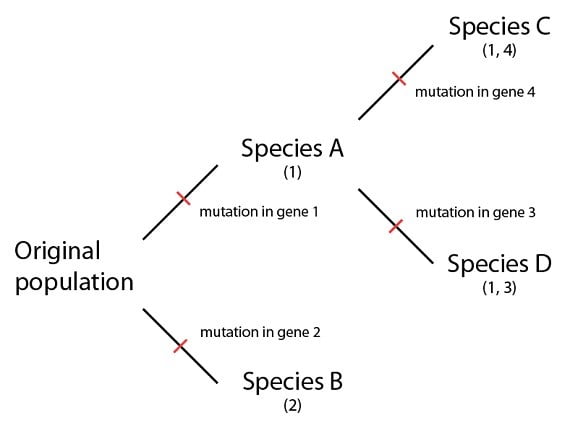
Si des événements de spéciation produisaient effectivement les espèces A-D à partir d’une population ancestrale commune, nous nous attendrions à voir leurs génomes montrer certaines caractéristiques lorsqu’ils sont comparés. D’abord, la séquence et la structure générales du génome devraient être hautement similaires – elles devraient être des versions du même livre, avec des chapitres, des paragraphes d’un texte partagé dans le même ordre. Ensuite, les différences entre elles devraient suivre un certain schéma. Les espèces C et D par exemple, devraient partager certaines caractéristiques qui seraient le résultat de leur ancêtre commun (l’espèce A) plus récent que celui qu’ils partagent avec l’espèce B. Dans ce simple diagramme, par exemple, les espèces C et D auraient une mutation identique dans le gène 1, et l’explication la plus parcimonieuse serait qu’elles l’ont toutes les deux héritées d’un ancêtre commun (l’espèce A). Cette explication est plus probable que celle qui avance que les deux espèces possèdent la même mutation, arrivées indépendamment l’une de l’autre, au même endroit, dans les deux génomes.
Plus généralement, l’hypothèse de l’ancêtre commun fait des prédictions spécifiques sur le schéma qu’on devrait observer en examinant les génomes. Dans le billet prochain, nous verrons à quel point ces prédictions tiennent la route lorsqu’on examine des données génomiques dans un groupe proposé d’espèces parentes.
48 Articles pour la série :
- 01-L’évolution expliquée : Introduction
- 02-L’Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l’homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l’humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour




