
Article 40 sur un total de 48 pour la série :

Photo credit: North Carolina School of Science and Mathematics (Creative Commons BY-NC-SA 2.0)
Dans les derniers billets de cette série, nous avons tracé les contours de l’évolution hominienne – les hominiens étant ces espèces (éteintes) qui sont plus proches de nous que ne le sont les chimpanzés, nos parents les plus proches qui soient encore en vie. Et comme nous l’avons vu, l’arbre généalogique de la famille hominienne s’est diversifié avec de nombreuses espèces qui coexistent dans le temps et dans l’espace.
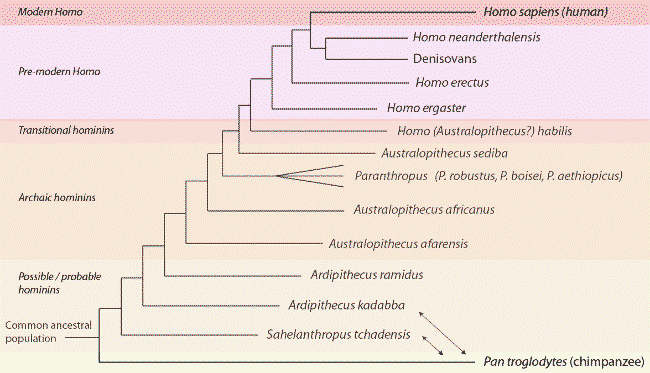
Phylogénie hominienne basée sur des preuves paléontologiques – et dans certains cas, génomiques – actuelles. Les séquences des génomes de l’humain, de Denisovan et de Néanderthal sont disponibles. Voir le billet précédent pour plus de détails.
Un fait s’avère particulièrement intéressant : c’est que ce patron d’espèces hominiennes contemporaines s’étend aux humains : nous avons coexisté avec d’autres espèces hominiennes pendant des milliers d’années. Le recouvrement géographique et temporel de l’humain et de Néanderthal par exemple est connu depuis longtemps. Ce qui demeurait inconnu, et hautement contentieux relevait de la question de la reproduction entre les humains et les groupes hominiens éteints. L’humain et le Néanderthal ont-ils eu des enfants ensemble ? Certains de ces enfants sont-ils restés avec les populations humaines devenant les ancêtres de certains humains modernes ? Il est quasiment impossible de traiter de telles questions en utilisant uniquement des restes fossiles (surtout puisque l’humain et le Néanderthal sont similaires). La génétique, en revanche, pourrait proposer une réponse définitive ; et les développements de la technologie génomique étant alors la clé pour résoudre le puzzle.
La naissance de la (paléo)génomique
Il est parfois difficile pour moi en tant que scientifique d’expliquer à mes amis à quel point la science qui séquence des génomes entiers a progressé cette dernière décennie. Lorsque j’étais étudiant à la fin des années 1990, même le génome de l’organisme modèle de choix (la mouche du vinaigre, Drosophila melanogaster) n’avait pas encore été entièrement séquencé. A cette époque, le projet du génome humain était en route, mais il était loin d’être complet. Le séquençage de génome était le truc de la « grande science » : de grands groupes bien subventionnés qui dépensaient des millions et des millions de dollars pour péniblement assembler les données, morceau par morceau.
Bien entendu, lorsque la communauté scientifique fait un tel investissement, elle gagne des dividendes qui vont bien au-delà des seules données obtenues ; nous gagnons aussi technologiquement, dans la mesure où nous devenons bien plus capables dans le domaine. En une décennie, le séquençage du génome, de coûteux et exclusif à la « grande science », est devenu une technique peu coûteuse, largement automatisée. La génomique n’est pas seulement devenue moins chère, elle est aussi devenue plus sensible : nous avons gagné la capacité d’assembler des données d’un génome à partir de plus petites quantités d’ADN.
Enfin, ces progrès nous ont permis d’isoler et de séquencer l’ADN d’espèces éteintes. Aussi étonnant que cela puisse sembler, dans les bonnes conditions, l’ADN peut persister à l’intérieur d’os et de dents des dizaines (voire des centaines) de milliers d’années. Etant donné notre intérêt pour l’évolution humaine, le séquençage de l’ADN de Néanderthal a été l’une des premières utilisations de cette technologie – et pour la première fois, il était possible de tester l’hypothèse d’une reproduction entre espèces avec des preuves génétiques.
Tous dans la famille, ou pas ?
Le premier ADN néanderthalien à être séquencé a été l’ADN mitochondrial, qui, comme vous vous en rappellerez sûrement, est hérité de la mère. Ces premières études ont montré que l’ADN mitochondrial néanderthalien n’avait pas été transmis à des populations humaines modernes. Ce qui ne rendait pas encore impossible la reproduction entre espèces dans la mesure où les lignages d’ADN mitochondrial peuvent facilement se perdre dans une population, et qu’un mâle néanderthalien se reproduisant n’introduirait pas d’ADN mitochondrial dans les populations humaines.
D’autres travaux de séquençage de l’ADN chromosomique néanderthalienont ensuite produit les résultats suivants : certaines populations humaines ont un ancêtre néanderthalien. Lorsque les humains ont migré hors d’Afrique il y a environ 50000 ans, ils ont rencontré des hommes de Néanderthal dans ce qu’on appelle aujourd’hui le Moyen Orient. Cette rencontre a compris des cas (relativement rares) de reproduction entre les deux groupes. De plus, nous savons qu’au moins certains des enfants des couples humain/néanderthal sont restés avec les populations humaines, et ont eu leurs propres enfants. De nos jours, on peut trouver des traces de l’ADN néanderthalien dans tous les humains modernes qui ne sont pas de descendance africaine subsaharienne. Puisque les Africains subsahariens descendent d’une population qui n’a pas rencontré de Néanderthal, ils ne possèdent pas cette variation d’ADN néanderthalienne caractéristique que l’on trouve dans d’autres populations humaines.
Plus de données, plus de preuves de reproduction entre espèces
Bien que le génome néanderthalien fût publié en 2010, nous avons fait depuis des progrès significatifs dans la compréhension des patrons de reproduction entre hominiens. La découverte d’un groupe jusque-là inconnu, Denisovan, que nous ne connaissons qu’à partir de quelques os de doigt et de dents, est un exemple d’un tel progrès. Ces maigres restes ont néanmoins permis un séquençage complet du génome denisovien, révélant qu’ils étaient des parents de Néanderthal. Le génome denisovien a également révélé qu’ils ont eux aussi contribué à l’ADN de certaines populations humaines modernes, spécifiquement des populations descendant des Mélanésiens.
Des travaux encore plus récents ont séquencé un génome néanderthalien de haute qualité. Bien que nous ayons depuis longtemps et de façon relativement abondante des restes des Néanderthal, la qualité de leur ADN est plutôt pauvre. La découverte de restes de Néanderthal dans le même lieu que celui qui a préservé l’ADN exceptionnel de Denisova a enfin donné un génome néanderthalien de très haute qualité, dont l’analyse a été publiée cette année. Ces nouvelles données ont permis plusieurs analyses plus robustes comparant les génomes humains, denisovien et néanderthalien. Ces analyses ont produit plusieurs résultats : des populations du continent asiatique possèdent aussi une petite quantité d’ADN denisovien ; les humains ont contribué à une variation de l’ADN néanderthalien ; Néanderthal et Denisova se sont reproduits entre eux, provoquant des variations d’ADN des deux côtés ; et (c’est peut-être ce qu’il y a de plus intéressant) Denisova possède des variations qui suggèrent qu’il s’est reproduit avec un autre groupe hominien archaïque, possiblement Homo erectus.
A partir de ces résultats, nous pouvons revoir l’arbre des espèces hominiennes et reconnaître que son patron n’est pas aussi simple que nous l’avions représenté. Les contours et les relations générales n’ont pas changé, mais au lieu de divisions claires entre des lignages séparés, nous devons plutôt imaginer le patron comme une toile, avec des échanges génétiques entre ce que nous avons auparavant représenté comme des espèces complètement distinctes. Alors que nous continuons à trouver de nouveaux restes hominiens et que nous séquençons leur ADN, il est probable que le patron devienne de plus en plus similaire à une toile, ajoutant des détails et de la complexité. Tout comme un historien découvre des anciens textes et des échanges entre des langues anciennes, de même nous pouvons désormais voir la fabrication faite à partir de différentes pièces de notre propre génome et de celui de nos proches parents.
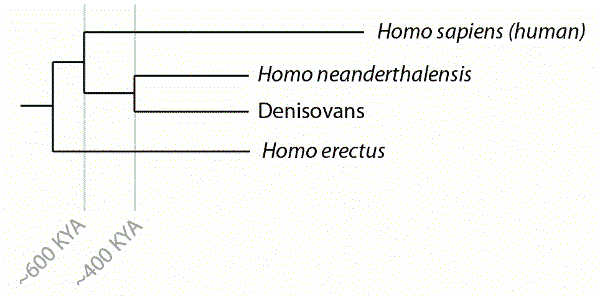
Relations hominiennes et dates approximatives de divergence des lignages conduisant au Néanderthal, à Denisova et à l’humain moderne. La paléogénomique a montré que ce patron a une structure plus proche d’une toile qu’à celle de branches d’un arbre, avec une reproduction limitée entre ces groupes.
Bien entendu, biologiquement parlant, ces découvertes compliquent la possibilité de trouver un “premier humain” et s’ajoutent aux problèmes habituels des changements progressifs d’une espèce au cours du temps. Les humains modernes sont une espèce, mais ont néanmoins des variations génomiques subtiles dérivées de rencontres régionales et d’échanges génétiques avec un certain nombre d’hominiens parents qui ont divergé de notre lignage et ont quitté l’Afrique avant notre expansion géographique. Aussi fascinant scientifiquement parlant que soient ces résultats, ces découvertes sont évidemment d’un intérêt théologique pour des chrétiens aussi ; une conversation qui commence à peine dans les cercles évangéliques, et qui prendra probablement du temps pour se développer.
Pour en savoir plus
- Neanderthals, Denisovans and Human Speciation”
- Were Adam and Eve historical figures?
- “Why the Church Needs Multiple Theories of Original Sin”
- James Kidder: “The Human Fossil Record
[p1]les garçons et les filles ont l’ADN mito de leur maman!
48 Articles pour la série :
- 01-L’évolution expliquée : Introduction
- 02-L’Evolution : Une théorie testée, pas une simple hypothèse !
- 03-Biogéographie
- 04-Une introduction à la variation, à la sélection naturelle et artificielle
- 05-Les chiens domestiques
- 06-Comment ça marche, la sélection naturelle ?
- 07-La sélection naturelle et le lignage humain.
- 08-Les bases de la variation héréditaire, première partie
- 09-Les bases de la variation héréditaire, deuxième partie.
- 10-De la variation à la spéciation (1)
- 11-De la variation à la spéciation (2)
- 12-De la variation à la spéciation 3
- 13-De la variation à la spéciation (4)
- 14-Les génomes sont comme des anciens textes (1)
- 15-Les génomes comparés aux textes anciens (2)
- 16-Les génomes comparés aux textes anciens (3): les origines de l’homme
- 17-Le génome comparé à un texte ancien (4)
- 18-Le génome comparé à un texte ancien (5): rattacher le tout ensemble.
- 19-Les arbres généalogiques des espèces, des gènes, et tri incomplet des lignées
- 20-Tri de lignage incomplet et taille d’une population ancestrale
- 21-Une introduction à l’homoplasie et à la convergence évolutive
- 22-Evolution convergente et homologie profonde.
- 23-Coévolution et la course à l’armement prédateur / proie
- 24-Le parasitisme, le mutualisme et la co-spéciation
- 25-Comprendre l’endosymbiose
- 26-La diversification cambrienne et la mise en place des plans d’organisation animaux. Première partie.
- 27-La diversification cambrienne et la mise en place des plans d’organisation animaux. 2e partie.
- 28-La mise en place des plans d’organisation des corps vertébrés, première partie.
- 29-La mise en place des plans d’organisation des corps vertébrés, deuxième partie.
- 30-La mise en place des plans d’organisation des vertébrés : Troisième partie.
- 31-La mise en place des plans d’organisation des vertébrés, quatrième partie.
- 32-Des reptiles aux mammifères.
- La révolution placentaire : deuxième partie.
- Du primate à l’humain, première partie.
- Du primate à l’humain, deuxième partie
- Du primate à l’humain (3)
- La paléontologie hominienne : une petite esquisse des preuves actuelles
- Devenir humain (1) : Eve mitochondriale et Adam Y-chromosomique
- Analogie entre évolution biologique et évolution du langage
- Devenir humain (3) : paléogénomiques et la toile emmêlée de la spéciation humaine
- 41-L’évolution, Partie 1: frontières scientifiques, abiogenèse et apologétique chrétienne
- 42-Aux frontières de l’évolution, Partie 2: l’abiogenèse et la question du naturalisme
- 43-Aux frontières de l’évolution, Partie 3 : l’hypothèse du monde à ARN
- 44-Aux frontières de l’évolution, partie 4 : Contingence versus convergence
- 45-Aux frontières de l’évolution, partie 5 : Contigence versus convergence dans l’expérience LTEE
- 46- L’évolution et le chrétien, première partie: Est-ce que l’évolution est un mécanisme sans but ?
- 47- L’évolution et le chrétien, deuxième partie: Une créature merveilleuse
- 48- L’évolution et le chrétien, partie 3 : Dire la vérité dans l’amour



